Тонический рефлекс на растяжение

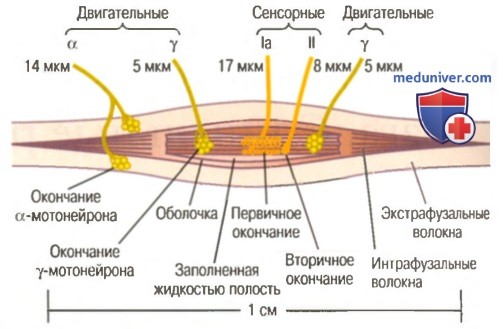
Реакция первичного окончания мышечного веретена. Рефлекс на растяжение мышцыа) Реакция первичного и вторичного окончаний на длину рецептора — статический ответ. Когда рецепторная часть мышечного веретена растягивается медленно, количество импульсов, передаваемых от первичных и вторичных окончаний, возрастает прямо пропорционально степени растяжения, и окончания продолжают передавать эти импульсы в течение нескольких минут. Этот эффект называют статическим ответом мышечного веретена. Это значит, что и первичные, и вторичные окончания продолжают передавать сигналы в течение нескольких минут, если само мышечное веретено остается растянутым.
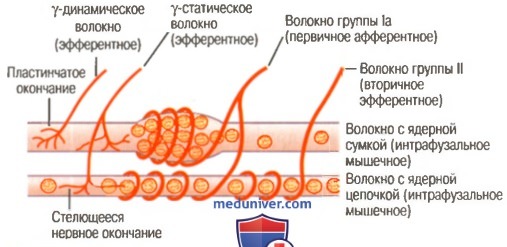
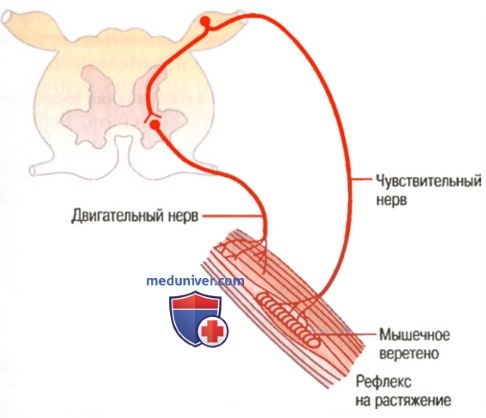
б) Реакция первичного окончания (но не вторичного) на скорость изменения длины рецептора — динамический ответ. Когда длина мышечного веретена изменяется внезапно, первичное окончание (но не вторичное) стимулируется особенно мощно. Это избыточное возбуждение первичного окончания называют динамическим ответному что означает чрезвычайно активную реакцию первичного окончания на высокую скорость изменения длины веретена. Даже когда длина веретена увеличивается лишь на долю микрометра и это увеличение осуществляется в течение доли секунды, первичный рецептор передает громадное количество дополнительных импульсов по большим сенсорным нервным волокнам диаметром 17 мкм, но только до тех пор, пока длина действительно увеличивается. Как только увеличение длины прекращается, этот дополнительный прирост импульсного разряда возвращается к гораздо более низкому уровню, чем статический разряд, все еще присутствующий в ответе. Наоборот, при укорочении веретена происходит прямо противоположное изменение сигнала. Таким образом, первичное окончание посылает чрезвычайно сильные, положительные или отрицательные, сигналы к спинному мозгу, информируя его о любом изменении длины мышечного веретена. в) Регуляция интенсивности статических и динамических ответов гамма-мотонейронами. Гамма-двигательные нервы к мышечному веретену можно разделить на два типа: гамма-динамические (гамма-d) и гамма-статические (гамма-s). Первые из них возбуждают главным образом интрафузальные волокна с ядерной сумкой, а вторые возбуждают в основном интрафузальные волокна с ядерной цепочкой. Когда гамма-d волокна возбуждают волокна с ядерной сумкой, динамический ответ мышечного веретена становится чрезвычайно усиленным, тогда как статический ответ почти не меняется. Наоборот, стимуляция гамма-s волокон, возбуждающих мышечные волокна с ядерной цепочкой, усиливает статический ответ, оказывая лишь незначительное влияние на динамический ответ. Непрерывный разряд мышечных веретен при нормальных условиях. В норме, особенно на фоне некоторой степени возбуждения гамма-эфферентных волокон, в сенсорных нервных волокнах мышечных веретен постоянно возникают импульсы. Растяжение мышечных веретен увеличивает частоту импульсов, тогда как укорочение веретен снижает ее. Таким образом, веретена могут посылать к спинному мозгу положительные сигналы, т.е. увеличенное количество импульсов, свидетельствующее о растяжении мышцы, или отрицательные сигналы, т.е. количество импульсов ниже нормы, что указывает на отсутствие растяжения мышцы. Рефлекс на растяжение мышцыСамым простым проявлением функции мышечных веретен является рефлекс на растяжение мышцы. Всякий раз, когда мышца внезапно растягивается, возбуждение веретен вызывает рефлекторное сокращение крупных мышечных волокон растянутой мышцы и тесно связанных с ней мышц-синергистов. а) Нервный контур рефлекса на растяжение. На рисунке ниже изображен основной контур рефлекса на растяжение мышечных веретен.
Видно, что проприоцептивное нервное волокно типа 1а, исходящее от мышечного веретена, входит в задний корешок спинного мозга. Затем ветвь этого волокна идет непосредственно к переднему рогу серого вещества спинного мозга и синаптически связывается с передними мотонейронами, которые посылают двигательные нервные волокна к той же мышце, от которой исходят волокна мышечных веретен. Таким образом, существует моносинаптический путь, позволяющий рефлекторному сигналу вернуться с кратчайшей задержкой назад к мышце после возбуждения веретена. Большинство волокон типа II от мышечных веретен заканчиваются на множестве вставочных нейронов серого вещества, и их аксоны проводят сигналы к передним мотонейронам с задержкой или выполняют другие функции. — Также рекомендуем «Динамические и статические рефлексы на растяжение. Демпфирующий механизм сокращения» Оглавление темы «Физиология спинальной рефлекторной деятельности»: |
Источник
Рефлекс на растяжение в клинике. Клонус мышечных волокон
а) Клиническое использование рефлекса на растяжение. При проведении клинического обследования пациента врач проверяет множество рефлексов на растяжение с целью определения степени фонового, или «тонического», возбуждения, передаваемого из головного мозга к спинному. Этот рефлекс вызывается следующим образом.
1. Коленный и другие мышечные рефлексы. В клинике для определения чувствительности рефлексов на растяжение обычно используют возбуждение коленного и других мышечных рефлексов. Для вызова коленного рефлекса ударяют неврологическим молоточком по сухожилию надколенника; это мгновенно растягивает четырехглавую мышцу бедра, что вызывает динамический рефлекс на растяжение, в результате которого нижняя конечность дергается вперед. На рисунке ниже представлена миограмма четырехглавой мышцы, зарегистрированная во время коленного рефлекса.
Миограммы четырехглавой мышцы бедра во время развития коленного рефлекса (вверху) и икроножной мышцы во время клонуса лодыжки (внизу)
Подобные рефлексы можно получить почти от любой мышцы тела при ударе либо по их сухожилию, либо по брюшку самой мышцы. Другими словами, все, что требуется для выявления динамического рефлекса на растяжение — это внезапное растяжение мышечных веретен.
Неврологи используют мышечные рефлексы для оценки степени облегчения спинномозговых центров. Когда большое количество облегчающих импульсов передается от верхних регионов центральной нервной системы в спинной мозг, мышечные рефлексы значительно усилены. Наоборот, если облегчающие импульсы подавлены или не действуют, мышечные рефлексы заметно ослаблены или отсутствуют.
Эти рефлексы чаще используют для определения наличия или отсутствия спастичности мышц при поражениях двигательных областей головного мозга или заболеваниях, возбуждающих бульборетикулярную облегчающую область ствола мозга. Обычно обширные поражения моторных областей коры большого мозга в отличие от нижерасположенных двигательных регуляторных областей (особенно поражения, связанные с инсультами или опухолями мозга) сопровождаются чрезмерно усиленными мышечными рефлексами на противоположной стороне тела.
б) Клонус — осцилляция мышечных рефлексов. При определенных условиях мышечные рефлексы могут осциллировать. Этот феномен называют клонусом (см. нижнюю миограмму на рисунке выше). Осцилляцию легче всего объяснить на примере клонуса лодыжки.
Если человек, стоящий на кончиках пальцев стоп (на цыпочках), внезапно встает на всю стопу, растягивая икроножную мышцу, импульсы от мышечных веретен передаются в спинной мозг. При этом растянутая мышца рефлекторно возбуждается и снова поднимает тело. Через долю секунды рефлекторное сокращение мышцы прекращается, и тело «падает» на стопу опять, таким образом растягивая веретена второй раз.
Снова динамический рефлекс растяжения поднимает тело, но он также прекращается через долю секунды, и тело падает снова, начиная новый цикл. Часто рефлекс растяжения икроножной мышцы продолжает осциллировать в течение длительного времени. Это и есть клонус.
Клонус обычно развивается только в том случае, если рефлекс растяжения сильно сенсибилизирован облегчающими импульсами из головного мозга. Например, у децеребрированного животного, у которого рефлексы растяжения сильно облегчены, клонус развивается легко. Для определения степени облегчения спинного мозга неврологи тестируют пациента на клонус путем внезапного растяжения мышцы и использования постоянной растягивающей ее силы. Если клонус развивается, степень облегчения, несомненно, высокая.
— Также рекомендуем «Сухожильный рефлекс Гольджи. Значение сухожильного рефлекса»
Оглавление темы «Физиология спинальной рефлекторной деятельности»:
1. Реакция первичного окончания мышечного веретена. Рефлекс на растяжение мышцы
2. Динамические и статические рефлексы на растяжение. Демпфирующий механизм сокращения
3. Гамма-эфферентная система мышечного сокращения. Стабилизация положения тела
4. Рефлекс на растяжение в клинике. Клонус мышечных волокон
5. Сухожильный рефлекс Гольджи. Значение сухожильного рефлекса
6. Функция мышечных веретен. Сгибательный рефлекс и механизм отдергивания
7. Перекрестный разгибательный рефлекс. Реципрокное торможение и иннервация
8. Позные и локомоторные рефлексы спинного мозга. Шагательные движения
9. Шагание на месте. Чесательный рефлекс и рефлексы мышечного спазма
10. Спинальные вегетативные рефлексы. Спинальный шок
Источник
The patellar tendon reflex is an example of the stretch reflex.
The stretch reflex (myotatic reflex), or more accurately «muscle stretch reflex», is a muscle contraction in response to stretching within the muscle. The reflex functions to maintain the muscle at a constant length. The term deep tendon reflex is often wrongfully used by many health workers and students to refer to this reflex. «Tendons have little to do with the response, other than being responsible for mechanically transmitting the sudden stretch from the reflex hammer to the muscle spindle. In addition, some muscles with stretch reflexes have no tendons (e.g., «jaw jerk» of the masseter muscle)».[1]
As an example of a spinal reflex, it results in a fast response that involves an afferent signal into the spinal cord and an efferent signal out to the muscle. The stretch reflex can be a monosynaptic reflex which provides automatic regulation of skeletal muscle length, whereby the signal entering the spinal cord arises from a change in muscle length or velocity. It can also include a polysynaptic component, as in the tonic stretch reflex.[2]
When a muscle lengthens, the muscle spindle is stretched and its nerve activity increases. This increases alpha motor neuron activity, causing the muscle fibers to contract and thus resist the stretching. A secondary set of neurons also causes the opposing muscle to relax.
Gamma motoneurons regulate how sensitive the stretch reflex is by tightening or relaxing the fibers within the spindle. There are several theories as to what may trigger gamma motoneurons to increase the reflex’s sensitivity. For example, alpha-gamma co-activation might keep the spindles taut when a muscle is contracted, preserving stretch reflex sensitivity even as the muscle fibers become shorter. Otherwise the spindles would become slack and the reflex would cease to function.
This reflex has the shortest latency of all spinal reflexes including the Golgi tendon reflex and reflexes mediated by pain and cutaneous receptors.[3]
Structures[edit]
The stretch reflex is accomplished through several different structures. In the muscle, there are muscle spindles, whose extrafusal muscle fibers lie parallel to the muscle and sense changes in length and velocity. The afferent sensory neuron is the structure that carries the signal from the muscle to the spinal cord. It carries this action potential to the dorsal root ganglion of the spinal cord. The efferent motor neuron is the structure that carries the signal from the spinal cord back to the muscle. It carries the action potential from the ventral root of the spinal cord to the muscle down the alpha motor neuron.[4] This synapses on the first structure discussed, the extrafusal fibers of the muscle spindle.
Examples[edit]
A person standing upright begins to lean to one side. The postural muscles that are closely connected to the vertebral column on the opposite side will stretch. The muscle spindles in those muscles will detect this stretching, and the stretched muscles will contract to correct posture.
Other examples (followed by involved spinal nerves) are responses to stretch created by a blow upon a muscle tendon:
- Jaw jerk reflex (CN V)
- Biceps reflex C5/C6
- Brachioradialis reflex C6
- Extensor digitorum reflex C6/C7
- Triceps reflex C6/C7
- Patellar reflex L2-L4 (knee-jerk)
- Ankle jerk reflex S1/S2
Another example is the group of sensory fibers in the calf muscle, which synapse with motor neurons innervating muscle fibers in the same muscle. A sudden stretch, such as tapping the Achilles’ tendon, causes a reflex contraction in the muscle as the spindles sense the stretch and send an action potential to the motor neurons which then cause the muscle to contract; this particular reflex causes a contraction in the soleus-gastrocnemius group of muscles. Like the patellar reflex, this reflex can be enhanced by the Jendrassik maneuver.
Spinal control[edit]
Spinal control of the stretch reflex means the signal travels between the muscle and spinal cord. The signal returns to the muscle from the same spinal cord segment as where it entered the spinal cord. This is the shortest distance for a reflex signal to travel, thus creating a fast response. These responses are often referred to short latency stretch reflexes.[5]
Supraspinal control[edit]
Supraspinal control of the stretch reflex means the signal travels above the spinal cord before traveling back down to the same segment it entered the spinal cord from. The responses from these pathways are often termed medium or long latency stretch reflexes, because the time course is longer due to distance it needs to travel.[6]
The central nervous system can influence the stretch reflex via the gamma motoneurons, which as described above control the sensitivity of the reflex.
Clinical significance[edit]
The knee jerk reflex is an example of the stretch reflex and it is used to determine the sensitivity of the stretch reflex.
Reflexes can be tested as part of a neurological examination, often if there is an injury to the central nervous system. To test the reflex, the muscle should be in a neutral position. The patient should flex the muscle being tested for the clinician to locate the tendon. After the muscle is relaxed, the clinician strikes the tendon. The response should be contraction of the muscle. If this is the knee jerk reflex, the clinician should observe a kick. The clinician rates the response.[7]
| Grade | Response | Significance |
|---|---|---|
| 0 | no response | always abnormal |
| 1+ | slight but definitely present response | may or may not be normal |
| 2+ | brisk physiologic response | normal |
| 3+ | very brisk response | may or may not be normal |
| 4+ | clonus | always abnormal |
The clasp-knife response is a stretch reflex with a rapid decrease in resistance when attempting to flex a joint. It is one of the characteristic responses of an upper motor neuron lesion.[9]
See also[edit]
- Proprioception
- Reflex
References[edit]
- ^ Evidence-Based Physical Diagnosis; McGee; Chapter 63. 2018
- ^ «Interaction between voluntary contraction and tonic stretch reflex transmission in normal and spastic patients». doi:10.1136/jnnp.35.6.853.
- ^ Spirduso, Waneen Wyrick (1978). «Hemispheric Lateralization and Orientation in Compensatory and Voluntary Movement». Information Processing in Motor Control and Learning. pp. 289–309. doi:10.1016/B978-0-12-665960-3.50019-0. ISBN 9780126659603.
- ^ Dolbow, James; Bordoni, Bruno (2019), «Neuroanatomy, Spinal Cord Myotatic Reflex», StatPearls, StatPearls Publishing, PMID 31869093, retrieved 2019-12-30
- ^ Feher, Joseph (2012). «Spinal Reflexes». Quantitative Human Physiology. pp. 332–340. doi:10.1016/B978-0-12-382163-8.00036-0. ISBN 9780123821638.
- ^ ELDRED E, GRANIT R, MERTON PA (1953). «Supraspinal control of the muscle spindles and its significance». J Physiol. 122 (3): 498–523. doi:10.1113/jphysiol.1953.sp005017. PMC 1366137. PMID 13118557.CS1 maint: multiple names: authors list (link)
- ^ Walkowski, A. D.; Munakomi, S. (2019). «StatPearls». PMID 31082072.
- ^ Walker, H. K.; Walker, H. K.; Hall, W. D.; Hurst, J. W. (1990). «Deep Tendon Reflexes». PMID 21250237. [1]
- ^ Ashby P, Mailis A, Hunter J (1987). «The evaluation of «spasticity»». Can J Neurol Sci. 14 (3 Suppl): 497–500. doi:10.1017/s0317167100037987. PMID 3315151.CS1 maint: multiple names: authors list (link)
External links[edit]
- Reflex,+Stretch at the US National Library of Medicine Medical Subject Headings (MeSH)
- «Stretch reflex» at Dorland’s Medical Dictionary
Источник
Тонические лабиринтные рефлексы. Рефлексы положения
Рефлексы положения (позы). Как известно, мышцы даже в покое находятся в некотором тоническом напряжении. Если отрезать мышцу от места прикрепления, она сразу же укоротится. Мышечный тонус обеспечивает фиксацию подвижных суставов и противодействует силе тяжести.
В отличие от быстрых, кратковременных и значительных по величине сокращений скелетных мышц, обозначаемых фазовыми движениями, тоническая деятельность осуществляется медленным изменением напряжения мышц, которое затем длительно удерживается. При этом происходит сокращение только незначительного количества мышечных волокон, остальные находятся в покое, затем активность переходит к другим волокнам. Поэтому мышечный тонус не обнаруживает признаков утомления.
Тонус мышц поддерживается главным образом благодаря миостатическим рефлексам (т. е. рефлексам на растяжение). Растяжение мышцы раздражает заложенные в ней проприоцепторы, и это дает начало тоническому рефлексу, препятствующему дальнейшему растяжению мышцы. Лабиринтиые рефлексы положения были изучены на децеребрированных животных (кошках) Магнусом и де Клейном.
Шеррингтон (Sherrington) показал, что если перерезать средний мозг между передним и задним четверохолмием, наступает децеребрациопная ригидность; у таких животных наблюдается крайне усиленный топус разгибателей. Этот экстензорный тонус настолько выражен, что животное может быть поставлено на вытянутые конечности. Если такое животное опрокинуть, оно не может восстановить утраченное равновесие, так как более сложная двигательная координация осуществляется только при сохранности таламуса (Thalamustier).
Импульсы, поддерживающие столь усиленный тонус, исходят из проприоцепторов шейных мышц п лабиринта (головного проприоцептора). Это доказывается перерезкой продолговатого мозга каудальнее вестибулярных ядер и шейного сегмента. После этого децеребрациопная ригидность проходит.
У децеребрированного животного очень демонстративно могут быть показаны шейные рефлексы, зависящие от изменения головы по отношению к туловищу.
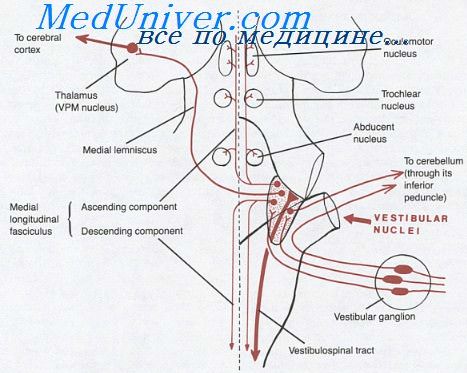
Если поднять или опустить голову животного или повернуть его влево или вправо пли дать наклон к правому или левому плечу,—тонус мускулатуры тела меняется совершенно закономерно. При подъеме головы, нап имер, тонус разгибателей передних конечностей увеличивается, а задних—уменьшается. Если опустить голову, то, наоборот, напряжние мышц задних конечностей увеличивается, а передних—уменьшается. При поворотах головы тонус мышц одной половины тела увеличивается, а другой—уменьшается.
Рефлексы эти исходят из проприоцепторов шейных мышц, которые при поворотах шеи раздражаются.
Классические опыты Магнуса и де Клейпа убедительно показали, что п лабиринты оказывают влияние на распределение тонуса мышц тела. Для исключения шейных рефлексов они загипсовали кошку таким образом, что голова ее была неподвижно фиксирована в отношении туловпща.
Поворачивая животное, как «куклу», они могли показывать, что любое положение головы в пространстве как бы «магическим» образом связано с распределением тонуса.
При расположении животного на спине (когда ротовая щель образует 45° с горизонтальной плоскостью) наблюдается наибольшее напряжение экстензоров. Наоборот, наименьший тонус они имеют при повороте животного на 180°, т. е. спиной вверх.
Такое же тоническое воздействие лабиринты оказывают на шейную мускулатуру, максимальное напряжение шейных мышц наблюдается при той же позиции головы, при которой экстензоры конечностей показывают максимальное напряжение.
При двусторонней экстирпации лабиринтов эти тонические рефлексы исчезают, так же как после отрыва отолитовых мембран центрифугированием. Этим доказано, что они исходят именно из отолитового прибора и не имеют отношения к ампулярному рецептору.
Как видно из вышеизложенного, отолитовые и шейные рефлексы работают содружественно, обеспечивая наиболее целесообразные позы животного.
Магнус и де Клейн считали, что тонические рефлексы исходят из утрикулюса, так как, согласно их гипотезе, максимальное раздражение отолитовый рецептор испытывает тогда, когда отолитовая мембрана отвисает, оказывая максимальную тягу за волоски невроэпителия. При отвисании отолитовой мембраны утрикулюса как раз и наблюдается максимум тонических рефлексов.
— Также рекомендуем «Компенсаторное движение глаз. Лабиринтные установочные рефлексы»
Оглавление темы «Отолитовый анализатор»:
1. Вегетативные рефлексы вестибулярного аппарата. Сенсорные реакции вестибулярной системы
2. Порог ощущения вестибулярного аппарата. Калорический раздражитель вестибулярного анализатора
3. Калорический нистагм. Характеристика калорической пробы
4. Механический раздражитель вестибулярного аппарата. Фистула вестибулярного аппарата
5. Электрический раздражитель вестибулярного аппарата. Отолитовый анализатор
6. Тонические лабиринтные рефлексы. Рефлексы положения
7. Компенсаторное движение глаз. Лабиринтные установочные рефлексы
8. Отолитовы мембраны. Функции отолитовой мембраны
9. Теория Квикса. Раздражители отолитового анализатора
10. Нистагм положения. Отолитовая реакция В.И. Воячека
Источник