Рецепторы мышц на растяжение

Специфические рецепторы поперечнополосатых мышц[править | править код]
Помимо свободных нервных окончаний (ноцицепторов) и рецепторов соединительнотканных футляров в мышцах и области мышечно-сухожильных соединений имеется два типа важных рецепторов глубокой чувствительности:
- мышечные веретена;
- сухожильные органы Гольджи.
Эти рецепторы относятся к группе проприорецепторов, которые играют важную роль в выполнении движений и поддержании позы. Проприорецепторы передают информацию о позе тела в пространстве и расположении конечностей относительно туловища (чувство положения). Кроме этого, они регистрируют движения в суставах и их скорость (чувство движения) и могут оценивать приложенное мышечное усилие (чувство силы). Эта информация далее направляется вместе с импульсами от органов чувств и равновесия в высшие центры моторной деятельности (в том числе в мозжечок). Эти импульсы передают основную информацию для контроля движений и коррекции поддержания позы и целенаправленных движений.
Мышечные веретена[править | править код]
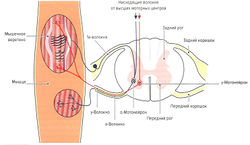
Рис. 1.6 а. Строение и функция мышечного веретена: кольцеспиральные нервные окончания 1а-волокон (Аа-волокна)
Мышечные веретена расположены по всей мышце. Их название обусловлено их веретенообразной формой. Количество мышечных веретен в мышце варьирует от 40 до 500 (Lindel, 2006). Как уже было отмечено, они лежат вдоль мышечных волокон (экстрафузальных волокон, вне веретена) и имеют 5-10 мм в длину и 0,2 мм в толщину (Kolster, 2006). Мышечные веретена состоят из соединительнотканной капсулы, поверхностной слой которой соответствует эндомизию, и 10-20 так называемых интрафузальных мышечных волокон (внутри веретена). Концы веретена прикрепляются к соединительнотканной оболочке мышцы или к сухожилию. Интрафузальные волокна только на концах имеют актиновые и миозиновые филаменты, позволяющие им сокращаться, а в центре волокно способно лишь к растяжению.
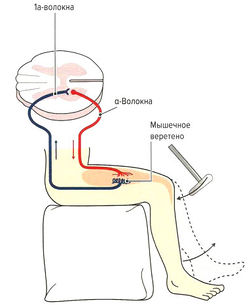
Рис. 1.6 б. Строение и функция мышечного веретена: моносинаптический проприоцептивный рефлекс
Несократительный растяжимый центр интрафузального волокна окружен толстым спиралевидным миелинизированным нервным окончанием типа 1а (Аа-волокна) (кольцеспиральное окончание) (рис. 1.6, а). Диаметр la-волокон составляет 10-20 мкм. Данные нервные волокна являются самыми быстрыми в организме человека — скорость распространения импульса составляет 60-120 м/с. Сократительные концы интрафузальных волокон иннервируются эфферентными у-волокнами (Ay-волокна, у-мотонейроны). Диаметр этих волокон составляет 4-8 мкм, а скорость передачи импульса — 20-50 м/с. Около 50 % всех мотонейронов передних рогов одного сегмента спинного мозга представлены у-мотонейронами (Cabri, 1999).
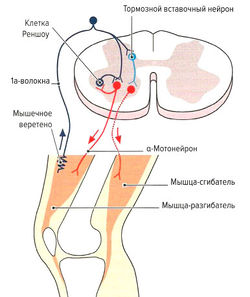
Рис. 1.6 в. Строение и функция мышечного веретена: реципрокное и возвратное ингибирование
Растяжение центральной части интрафузальных волокон приводит к раздражению кольцеспиральных нервных окончаний мышечного веретена (рецептор растяжения). На конце афферентных волокон возникает потенциал действия, который затем передается в ЦНС. Центральная часть веретена растягивается либо при растяжении всей мышцы, либо при напряжении сократительных концов интрафузальных волокон при активации у-мотонейронов. При одновременном действии этих двух факторов возможно как взаимное усиление, так и уменьшение их действия. Таким образом, мышечные веретена способны регистрировать как изменение длины мышцы, так и скорость ее изменения, а также косвенно ее изменять. Действуя через различные контуры регуляции, мышечные веретена выполняют несколько функций.
- Защита мышцы от перерастяжения за счет рефлекса растяжения (моносинаптический рефлекс). Функцией моносинаптического рефлекса растяжения является поддержание мышечной длины. Любое растяжение мышцы, например при ударе по сухожилию или мышце (рис. 1.6, б) или при изменении позы туловища, влияет на натяжение мышечных веретен. В результате резко повышается частота импульсации по 1а-афферентным нервным волокнам, идущим через задние спинномозговые корешки в серое вещество спинного мозга и через один синапс (моносинаптически) связываются с а-мотонейронами этой же мышцы (проприоцептивный рефлекс) (рис. 1.6, б). а-Мотонейроны по а-волокнам вызывают сокращение экстрафузальных волокон натянутой мышцы. Поскольку рецептор и эффектор находятся в одном и том же органе, скорость данного рефлекса очень высока (около 30 мс). Непроизвольные изменения длины мышцы могут сразу корректироваться на уровне спинного мозга без вовлечения высших моторных центров. Одновременно с этим через тормозной вставочный нейрон (ингибирующий la-нейрон) происходит ингибирование мотонейронов мышц-антагонистов (полисинаптический рефлекс) (реципрокное ингибирование антагонистов) (рис. 1.6, в). Благодаря этому растянутая мышца не преодолевает сопротивление антагонистов и предотвращается аналогичная реакция антагонистов при их растяжении (Weiss, 2000). После сокращения растянутой мышцы импульсация от la-волокон снижается. Дополнительно вследствие активации а-мотонейронов через коллатеральные связи и тормозные вставочные нейроны (клетки Реншоу) происходит обратное ингибирование самих а-мотонейронов (рис. 1.6, в). Эта отрицательная связь останавливает распространение сигнала и обеспечивает кратковременность сокращения мышцы, поэтому, несмотря на постоянные внешние раздражители (растяжение мышцы), возможно поддержание ее постоянной длины. Кроме этого, остановка рефлекторного ответа обеспечивается сухожильными органами Гольджи, которые активируются при мышечном сокращении и последующем растяжении сухожилия (аутогенное ингибирование) (см. рис. 1.7, б). В заключение необходимо отметить, что возбудимость мотонейронов контролируется высшими центрами головного мозга. При повреждении этих центров или отходящих от них нервных путей (например, при параплегии) развивается аномальное повышение рефлексов (см. рис. 1.6, а).
- Регуляция активного мышечного тонуса. Высшие моторные центры в головном мозге через так называемые экстрапирамидные пути активируют у-мотонейроны и через мышечные веретена регулируют длину мышц и, следовательно, активный мышечный тонус. Активация у-мотонейронов вызывает напряжение сократительных концов интрафузальных волокон, что растягивает центр мышечных веретен и приводит к активации la-волокон. Как уже было отмечено, импульс затем передается через задние рога в передние рога спинного мозга и возбуждает а-мотонейроны. а-Мотонейроны, в свою очередь, вызывают сокращение волокон скелетных
мышц, что опосредованно уменьшает натяжение центральной части веретен. Эта рефлекторная реакция продолжается до тех пор, пока 1а-волокна не будут регистрировать натяжение (у-петля). При нормальной позе (сидя, стоя или при ходьбе) в большинстве мышц, особенно в работающих против силы тяжести, регистрируется постоянная невысокая активность афферентных волокон мышечных веретен, что позволяет поддерживать определенный активный мышечный тонус. Мышцы, действующие против силы тяжести, например мышца, разгибающая позвоночник, в положении стоя находятся в постоянном небольшом натяжении, т. к. туловище под влиянием силы тяжести наклоняется несколько кпереди. За счет постоянной активации у-мотонейронов достигается дополнительное растяжение средней части мышечных веретен, что вызывает активацию высших моторных центров для поддержания мышечного тонуса. Благодаря постоянной фоновой афферентной импульсации во время обычных движений мышечные веретена могут воспринимать как укорочение, так и растяжение мышцы. При усилении или уменьшении активности у-мотонейронов изменяется чувствительность мышечных веретен и регулируется их активность при конкретных позе и движении. Таким образом, у-мотонейроны устанавливают в каждой конкретной ситуации оптимальную рабочую область мышечных веретен, и они могут регистрировать мышечную длину и скорость ее изменения. Это позволяет устанавливать и поддерживать постоянный активный мышечный тонус, а следовательно, поддерживать равновесие и положение суставов и определенную позу (чувство позы и опоры).
- Выполнение точных движений и регуляция тонкой моторики. Возможность регулировать чувствительность мышечных веретен, особенно вместе с произвольными движениями, играет большую роль. При произвольных движениях, например при разгибании в коленном суставе, происходит помимо активации а-мотонейронов и повышение активности у-мотонейронов. За счет а-у-коактивации устанавливается оптимальная длина мышечных волокон, чтобы поддерживать их чувствительность во время всего движения. При сокращении регистрируется растяжение мышцы и становятся возможными очень точные движения. Крохме того, в научной среде продолжаются дискуссии, являются ли точные движения конечностями исключительно результатом изменения у-активности. Супраспинальные нервные центры могут изменять у-активность в у-петле (изменение частоты импульсации по 1а-волокнам) и таким образом изменять активность связанных с ними а-мотонейронов (Weiss, 2005).
![]() Запомните: Чем больше мышечных веретен в мышце, тем более точные и дифференцированные движения возможны при ее сокращении. Например, в мышцах кисти и глазного яблока находится большое количество проприорецепторов (Kolster, 2005).
Запомните: Чем больше мышечных веретен в мышце, тем более точные и дифференцированные движения возможны при ее сокращении. Например, в мышцах кисти и глазного яблока находится большое количество проприорецепторов (Kolster, 2005).
Сухожильные органы Гольджи[править | править код]
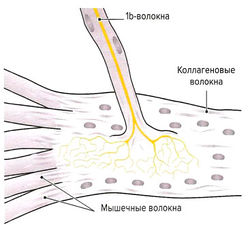
Рис. 1.7 а. Сухожильные органы Гольджи
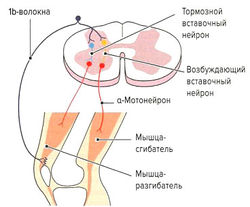
Рис. 1.7 б. Принцип аутогенного торможения
Помимо мышечных веретен, которые регулируют длину мышцы, в поперечнополосатой мышечной ткани находятся сухожильные органы Гольджи, регулирующие напряжение мышцы. Они располагаются вдоль экстрафузальных мышечных волокон в области мышечно-сухожильных соединений (рис. 1.7, а) и вплетаются в коллагеновые волокна сухожилия. Органы Гольджи состоят из множества разветвляющихся терминальных окончаний аксона lb-волокон (Аа-волокна), окруженных общей соединительнотканной капсулой. Эти рецепторы имеют в длину около 100 мкм и прикрепляются к 10-15 сухожильным волокнам (Cabri, 1999). При сокращении мышцы коллагеновые волокна натягиваются и сближаются. Расположенные между коллагеновыми волокнами нервные окончания сдавливаются и генерируют потенциал действия, который в виде нервного импульса передается по lb-волокнам в ЦНС. Адекватньш раздражителем для сухожильных органов Гольджи является изменение натяжения сухожилия, что происходит как при укорочении, так и при растяжении мышцы. Порог чувствительности этих рецепторов выше, чем у мышечных веретен. В отличие от моносинаптической дуги веретен, на уровне спинного мозга переключение импульса на а-мотонейрон идет через два или три синапса. Через тормозные вставочные нейроны (ди- или трисинаптически) происходит ингибирование а-мотонейронов заинтересованной мышцы аутогенное торможение). Одновременно с этим органы Гольджи через два синапса активируют через возбуждающие вставочные нейроны а-мотонейроны мышцы-антагониста (рис. 1.7, б). Оба этих механизма защищают мышцу от избыточного натяжения (защита от перегрузки).
Читайте также[править | править код]
- Поперечно-полосатые мышцы
- Кровоснабжение скелетных мышц
- Иннервация поперечно-полосатых мышц
- Cтроение и функции нейрона
- Синаптическая передача
- Нервно-мышечная передача
- Двигательная единица мыщцы
- Мышечно-сухожильное соединение
- Сухожилие
- Костно-сухожильное соединение
- Типы мышц
- Типы мышечных волокон
- Функции поперечно-полосатых скелетных мышц
- Гладкие мышцы
- Сердце
Источник
Во второй лекции по дисциплине «Биомеханика мышц» для студентов НГУ им. П.Ф.Лесгафта описывается состав и строение мышечных веретен — рецепторов, расположенных в мышцах и информирующих нервную систему о текущей длине мышцы и скорости ее сокращения, и сухожильных рецепторах Гольджи, которые информируют нервную систему о степени возбуждения мышцы. Также в лекции описываются уровни построения движений по Бернштейну и их взаимосвязь с функционированием рецепторами опорно-двигательного аппарата.
Лекция 2
Функционирование рецепторного аппарата мышц
2.1. Рецепторы опорно-двигательного аппарата человека
При изучении анатомии и физиологии (А.С. Солодков, Е.Б. Сологуб, 2001) вы изучали двигательную сенсорную систему. Одним из отделов этой системы являются проприорецепторы, расположенные в мышцах, сухожилиях и суставных сумках. В мышцах расположены мышечные веретена, в сухожилиях – сухожильные органы Гольджи. В суставных сумках расположены рецепторы суставов.
Мышечные веретена
Еще в XIX веке В. Кюне обнаружил в скелетных мышцах структуры, напоминающие веретено. Затем, в начале XX века Нобелевский лауреат Чарльз Скотт Шеррингтон показал, что эти структуры служат чувствительными рецепторами. Мышечные веретена рассеяны по всем скелетным мышцам. Концы их обычно прикрепляются к мышечным волокнам параллельно. Каждое веретено покрыто капсулой, которая расширятся в центре и образует ядерную сумку. Внутри веретена содержатся интрафузальные мышечные волокна. Эти волокна в 2-3 раза тоньше обычных (экстрафузальных) волокон скелетных мышц.
Интрафузальные волокна подразделяются на два типа:
- Длинные и толстые (диаметр 20-25 мкм)[1], которые информируют ЦНС о динамическом компоненте движения – скорости изменения длины мышцы. Таких волокон в мышечном веретене не более двух.
- Короткие и тонкие (диаметр 10–12 мкм), которые информируют ЦНС о статическом компоненте движения – текущей длине мышцы. Таких волокон в мышечном веретене от 2 до 12.
Рекомендую обратить внимание на учебные пособия «Биомеханика мышц» и «Гипертрофия скелетных мышц человека«
Рецепторы Гольджи
Нервно-сухожильные веретена (рецепторы Гольджи) открыл в 1903 году Камилло Гольджи. Впоследствии за эти исследования ему была присуждена Нобелевская премия. Рецепторы Гольджи располагаются в месте перехода мышечных волокон в сухожилия. Их длина составляет 0,5-1,0 мм, а диаметр – 0,1-0,2 мм. Отдельный нервный аксон несет афферентные импульсы в спинной мозг и называется аксоном Ib. Он начинается в виде веточек, проходящих между коллагеновыми волокнами сухожилия. Когда мышечные волокна сокращаются, коллагеновые волокна сухожилия натягиваются и сжимают нервные веточки, которые начинают импульсировать (рис.2.1). Таким образом, в результате последовательного крепления сухожильных органов к мышечным волокнам они возбуждаются при укорочении возбужденной мышцы. Сухожильные рецепторы возбуждаются в 1,5 – 8 раз более эффективно при мышечном сокращении, нежели при пассивном растяжении.
Рис. 2.1. Строение сухожильного органа Гольджи (А.Дж. Мак-Комас, 2001)
Рецепторы суставов
Суставные рецепторы подразделяются на несколько типов в зависимости от их реакции на амплитуду, скорость и направление движения в суставе.
Тельца Руффини находятся в капсуле сустава и воспринимают направление и скорость изменения межзвенного угла. Частота их импульсации возрастает с увеличением скорости изменения суставного угла.
Тельца Паччини посылают в ЦНС информацию о положении отдельных частей тела в пространстве и относительно друг друга. Эти рецепторы посылают в ЦНС информацию о значениях межзвенных углов, то есть о положении сустава. Их импульсация продолжается в течение всего периода сохранения межзвенного угла, и она тем больше, чем больше изменения угла.
2.2. Уровни построения движений и рецепторы опорно-двигательного аппарата
В 1947 году была опубликована книга русского ученого Н.А. Бернштейна «О построении движений». В этой книге он изложил оригинальную теорию управления двигательными действиями. В основе этой теории лежало понятие уровней построения движений. Он писал: «Каждая двигательная задача находит себе в зависимости от своего содержания и смысловой структуры тот или иной уровень. Иначе говоря, тот или иной сензорный синтез, который наиболее адекватен по качеству и составу образующих его афферентаций и по принципу их синтетического объединения требующемуся решению этой задачи. Этот уровень и определяется как ведущий уровень для данного движения» (Н.А.Бернштейн, 1991, С.41). По концепции Н.А. Бернштейна ни одно движение не обслуживается только одним уровнем, но осознается только ведущий уровень. По классификации Н.А.Бернштейна, при управлении движениями может быть задействовано пять уровней. Проприорецепторы опорно-двигательного аппарата человека составляют основу двух нижних уровней: уровня А и В.
Афферентация уровня А основана на импульсации мышечных веретен (длина и скорость сокращения мышцы) и рецепторов Гольджи (уровень возбуждения мышцы при ее укорочении). Эта информация очень слабо осознается ЦНС, то есть, по гипотезе Н.А.Бернштейна, этот уровень почти никогда не бывает ведущим.
Афферентация уровня В опирается на информацию, поступающую от суставных рецепторов. Это уровень выступает как ведущий в ряде физических упражнений, таких как наклоны тела вперед и назад, а также циклические движения. Сигналы от суставных рецепторов хорошо осознаются.
Рекомендуемая литература
1. Самсонова, А.В. Гипертрофия скелетных мышц человека: учебное пособие /А.В.Самсонова, СПб: Кинетика.- 159 с., ил.
2. Самсонова А.В. Биомеханика мышц: учебно-методическое пособие /А.В.Самсонова, Е.Н. Комиссарова / Под ред А.В.Самсоновой .- СПб., 2008.- 217 с.
3. Самсонова А.В. Моторная и сенсорная функции мышц в биомеханике локомоций: монография /А.В.Самсонова; СПбГУФК им. П.Ф.Лесгафта, 2007.- 152 с.
Источник
В активной части опорно двигательного аппарата имеются проприорецепторы. Существует два типа проприорецепторов – нервно-мышечные веретёна, которые реагируют на изменение длины мышцы, а также сухожильные органы Гольджи, реагирующие на изменение напряжения, то есть на силу сокращения мышечных волокон (Рисунок 1).

Рисунок 1. Проприорецепторы: нервно-мышечное веретено и сухожильный орган Гольджи
Нервно-мышечные веретёна. Нервно-мышечные веретена расположены в толще скелетных мышц между мышечными волокнами. Они представляют собой веретеновидные структуры, покрытые соединительнотканной капсулой. Длина веретена составляет 4 – 7 (до 10) мм, диаметр – 100 мкм. Под капсулой внутри веретена находятся видоизменённые мышечные волокна, которые называются интрафузальными. Эти волокна имеют ряд отличий от обычных, экстрафузальных, мышечных волокон. Во-первых, интрафузальное волокно имеет меньший диаметр (15 – 30 мкм) по сравнению с экстрафузальным (50 – 100 мкм). Во-вторых, в центральной («экваториальной») части интрафузального волокна нет сократительных элементов, поэтому к сокращению способны только периферические части. Поскольку концы интрафузального волокна фиксированы к соединительнотканной капсуле, покрывающей веретено, при укорочении периферических отделов волокна его центральная часть растягивается. Интрафузальные волокна расположены параллельно эктсрафузальным. По строению интрафузальные волокна делятся на два типа – с ядерной сумкой и с ядерной цепочкой (Рисунок 2). В свою очередь, волокна с ядерной сумкой по функции по особенностям иннервации подразделяются на статические и динамические. Для волокон с ядерной сумкой характерно наличие расширенной центральной части, в которой ядра собраны в скопления. Волокна с ядерной цепочкой примерно вдвое тоньше волокон с ядерной сумкой, ядра в их центральной части расположены в ряд. В каждом веретене содержится 1 – 3 волокна с ядерной сумкой и 5 – 7 волокон с ядерной цепочкой.

Рисунок 2. Нервно-мышечное веретено
Интрафузальные волокна получают как афферентную, так и эфферентную иннервацию. Выделяют два типа афферентных окончаний – первичные и вторичные. Первичные окончания называют аннулоспиральными, они представляют собой спираль, окружающую центральную часть всех интрафузальных волокон. От первичных окончаний отходят волокна типа Ia, которые соответствуют типу Aα по Гассеру-Эрлангеру. Это самые толстые волокна (~17 мкм) с самой высокой скоростью проведения возбуждения (70 – 12 м/с). Каждое нервно-мышечное веретено иннервируется только одним волокном Ia. Другими словами одно афферентное нервное волокно Ia получает импульсацию со всех первичных окончаний в иннервируемом им веретене. Вторичные окончания имеют форму спирали, но могут также образовывать разветвлённую структуру. Они оплетают центральную часть интрафузальных волокон с ядерной цепочкой, а также статических волокон с ядерной сумкой, но их не бывает в динамических волокнах с ядерной сумкой. От вторичных окончаний отходят волокна типа II, соответствующие типу Аβ по Гассеру-Эрлангеру; диаметр этих волокон составляет ~8 мкм, они проводят возбуждение со скоростью 40 – 70 м/с (Рисунок 3).

Рисунок 3. Схема нервно-мышечного веретена
Как первичные, так и вторичные нервные окончания чувствительны к изменению длины интрафузальных волокон, однако характер ответной реакции различается. При медленном растяжении центральной части интрафузальных волокон импульсация появляется как в первичных, так и во вторичных рецепторах. Частота импульсов пропорциональная степени растяжения волокна. Такой ответ афферентных окончаний называют статическим. Однако при быстром внезапном растяжении волокна в первичных (но не вторичных) окончаниях отмечается мощный всплеск импульсации – динамический ответ. Следовательно, волокна Ia передают информацию как об абсолютной длине интрафузальных волокон (то есть о степени растяжения), так о скорости её изменения, тогда как волокна II – только об абсолютной длине. Следует понимать, что растяжение центральной части интрафузальных волокон может быть связано как с удлинением всей мышцы (интрафузальные волокна расположены параллельно экстарфузальным), так и при сокращении периферических отделов волокна (Рисунок 3).

Рисунок 4. Статический и динамический ответы рецепторов нервно-мышечных веретён
Эфферентная иннервация интрафузальных волокон представлена нервными волокнами типа Aγ, которые являются аксонами γ-мотонейронов спинного мозга. Диаметр этих волокон составляет ~5 мкм, скорость проведения возбуждения в них – 15 – 40 м/с. Окончания Aγ волокон подходят к периферическим отделам интрафузальных волокон, при возбуждении γ-мотонейронов концевые участки интрафузальных волокон сокращаются, в результате чего их центральные отделы растягиваются. Выделяют γ-динамические и γстатические волокна, γ-динамические волокна иннервируют динамичекие волокна с ядерной сумкой, их активация повышает динамическую чувствительность окончаний Iа типа.
γ-статические волокна иннервируют волокна с ядерной цепочкой и статические волокна с ядерной сумкой, их возбуждение повышает уровень активности как первичных, так и вторичных окончаний (см. ниже).
Миотатический рефлекс (рефлекс растяжения)
В ответ на увеличение длины мышцы рецепторы нервно-мышечных веретён возбуждаются, и импульсация по волокнам Ia (и при быстром, и при медленном растяжении) или II (при медленном растяжении) поступает в спинной мозг. Войдя в серое вещество спинного мозга, волокно афферентного нейрона разветвляется и образует возбуждающие синапсы практически со всеми α-моторными нейронами, иннервирующими гомонимную (то есть эту же) мышцу. Возбуждение α-моторных нейронов вызывает сокращение экстрафузальных волокон, и мышца укорачивается. Следует отметить, что коллатерали афферентных нейронов переключаются также на вставочные тормозные нейроны, которые угнетают α-мотонейроны мышц антагонистов. Таким образом, растяжение мышцы вызывает её рефлекторное укорочение, при этом мышцы антагонисты расслабляются. Описанный рефлекс с рецепторов нервно-мышечных веретён называется миотатическим, или рефлексом на растяжение (Рисунок 4). Миотатический рефлекс является моносинаптическим, поэтому имеет очень небольшой латентный период (~ 0,9 мс).

Рисунок 5. Миотатический рефлекс с нервно-мышечных веретён
Различают фазический (динамический) и тонический миотатический рефлекс. Фазический рефлекс запускается с первичных нервных окончаний при быстром растяжении мышцы, например, при ударе по сухожилию неврологическим молоточком или при появлении внезапного препятствия совершаемому движению. Тонический рефлекс запускается с вторичных нервных окончаний при медленном (или постоянном) растяжении мышечных волокон; он имеет значение для поддержания вертикальной позы. Когда тело отклоняется от правильного положения, мышцы, участвующие в поддержании позы, растягиваются. В результате возбуждаются как первичные, так и вторичные окончания нервно-мышечных веретён, запускается миотатический рефлекс, и мышцы сокращаются; за счёт их сокращения тело возвращается в правильное положение. Когда у длительно стоящего человека (например, солдата в стойке «смирно») из-за утомления сгибаются ноги в коленных суставах, проприорецепторы четырёхглавых мышц реагируют на растяжение, и ноги снова выпрямляются. Постоянное стимулирующее влияние на α-мотонейроны со стороны афферентных волокон Ia и II типов вносит вклад в поддержание мышечного тонуса.
Функции γ-моторных нейронов. Понятие о γ-петле.
Как уже упоминалось, интрафузальные волокна получают эфферентную иннервацию со стороны γ-моторных нейронов. При возбуждении γ-мотонейронов происходит сокращение периферических (концевых) отделов интрафузальных волокон, в результате их центральная часть растягивается. При этом первичные и вторичные нервные окончания активируются, импульсация от них поступает в спинной мозг, и возбуждаются α-моторные нейроны. Другими словами, запускается миотатический рефлекс. Такой механизм (активация рефлекса растяжения при возбуждении γ-мотонейронов) называется γ-петлёй. Наличие γ-петли обеспечивает сохранение чувствительности рецепторов нервномышечных веретён при разной длине мышцы. Во время сокращения мышцы её длина уменьшается, соответственно должны укорачиваться и интрафузальные волокна. В результате импульсация с рецепторов нервно-мышечных веретён должна была бы прекратиться, и рецепторы потеряли бы способность реагировать на изменение длины мышцы. Напротив, при расслаблении мышцы её длина увеличивается, и интрафузальные волокна могут быть чрезмерно растянуты. В таких условиях рецепторы будут генерировать импульсы с максимальной частотой и, следовательно, также потеряют способность реагировать на изменение длины. Однако в реальных условиях имеет место α-γ-коактивация, то есть α- и γмотонейроны возбуждаются одновременно. В результате сокращаются как экстра-, так и интрафузальные волокна. Сокращение экстрафузальных волокон приводит к укорочению мышцы, а сокращение периферических отделов интрафузальных волокон – к растяжению их центральной части. Следовательно, длина центральной части не меняется, поскольку её укорочение вследствие сокращения экстрафузальных волокон компенсируется растяжением из-за сокращения периферических отделов интрафузальных волокон. И наоборот – расслабление всей мышцы сопровождается расслаблением и интрафузальных волокон, при этом растяжение центральной части уменьшается. Таким образом, за счёт α-γ-коактивации рецепторы нервно-мышечных веретён сохраняют чувствительность (и, следовательно, передают информацию о длине мышцы в ЦНС) как при сокращении мышцы, так и при её расслаблении (Рисунок 6). На γ-мотонейронах заканчиваются нисходящие пути из различных отделов головного мозга. Меняя активность γ-мотонейронов, мозговые центры влияют на чувствительность рецепторов нервно-мышечных веретён и, следовательно, на порог рефлекса растяжения. Сухожильные органы Гольджи. Обратный миотатический рефлекс Кроме нервно-мышечных веретён в активной части опорно-двигательного аппарата имеется ещё один тип проприрецепторов – сухожильные органы Гольджи. Сухожильные органы представляют собой сеть коллагеновых нитей, окружённую тонкой соединительнотканной капсулой. Диаметр сухожильного органа Гольджи составляет 100 мкм, длина – 1 мм. Коллагеновые нити внутри органа оплетённых нервными окончаниями, от которых отходят нервные волокна типа Ib. эти нервные волокна, как и Ia, соответствуют типу Aα по Гассеру-Эрлангеру. Сухожильные органы Гольджи расположены в месте прикрепления мышечных волокон к сухожильным, при этом каждый сухожильный орган связан с 3 – 25 (в среднем – с 10 – 15) мышечными волокнами (Рисунок 7). Таким образом, в отличии от нервно-мышечных веретён, которые ориентированы параллельно мышечным
волокнам, сухожильные органы по отношению к мышечным волокнам расположены последовательно.

Рисунок 6. Функция γ-петли
Следовательно, сухожильные органы реагируют не на изменение длины мышцы, а на развиваемое ей усилие, другими словами, на степень её напряжения. Афферентные нервные волокна от сухожильных органов Гольджи следуют в спинной мозг, где, в отличие от чувствительных волокон с нервно-мышечных веретён, переключаются на вставочные тормозные нейроны. Вставочные нейроны, в свою очередь, образуют синаптические контакты с α-мотонейронами гомонимной мышцы. Таким образом, активация сухожильных органов приводит к расслаблению мышцы, к которой крепится данное сухожилие. Описанный рефлекс называется обратным рефлексом растяжения (обратным миотатическим рефлексом) (Рисунок 8). Этот рефлекс, по-видимому, имеет две функции. Во-первых, он предупреждает чрезмерное напряжение скелетных мышц, которое может привести к тяжёлым травмам (разрыву мышцы, отрыву сухожилия от кости или от мышцы). Во-вторых, обратный миотатический рефлекс обеспечивает равномерность силы сокращения разных мышечных волокон в пределах одной мышцы. Те волокна, которые напряжены слишком сильно, расслабляются за счёт усиления тормозных влияний со стороны спинного мозга. Напряжение волокон, которые сокращаются недостаточно сильно, напротив, усиливается за счёт уменьшения тормозного воздействия.

Рисунок 7. Сухожильный орган Гольджи
Проприорецепторы посылают в головной мозг информацию о состоянии скелетно-мышечной системы Импульсация с проприорецепторов поступает не только в спинной мозг, но и по коллатералям афферентных волокон в различные отделы головного мозга. В частности, импульсы с нервно-мышечных веретён и сухожильных органов следуют по спинномозжечковым трактам в мозжечок, а также по системе задних столбов (тонкий и клиновидный пучок) в продолговатый мозг, затем – в таламус, а оттуда – в кору больших полушарий. Таким образом, мозжечок и кора большого мозга постоянно получают информацию о состоянии скелетно-мышечной системы, а именно о длине мышц и о степени их напряжения. Эта информация является необходимой как для регуляции произвольных движений и позы, так и для осознания положения частей тела в пространстве и друг относительно друга.

Рисунок 8. Обратный миотатический рефлекс с сухожильных органов Гольджи
Источник