Рецептор растяжения в мышце это

Специфические рецепторы поперечнополосатых мышц[править | править код]
Помимо свободных нервных окончаний (ноцицепторов) и рецепторов соединительнотканных футляров в мышцах и области мышечно-сухожильных соединений имеется два типа важных рецепторов глубокой чувствительности:
- мышечные веретена;
- сухожильные органы Гольджи.
Эти рецепторы относятся к группе проприорецепторов, которые играют важную роль в выполнении движений и поддержании позы. Проприорецепторы передают информацию о позе тела в пространстве и расположении конечностей относительно туловища (чувство положения). Кроме этого, они регистрируют движения в суставах и их скорость (чувство движения) и могут оценивать приложенное мышечное усилие (чувство силы). Эта информация далее направляется вместе с импульсами от органов чувств и равновесия в высшие центры моторной деятельности (в том числе в мозжечок). Эти импульсы передают основную информацию для контроля движений и коррекции поддержания позы и целенаправленных движений.
Мышечные веретена[править | править код]
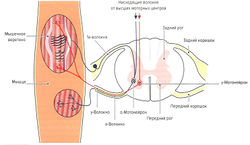
Рис. 1.6 а. Строение и функция мышечного веретена: кольцеспиральные нервные окончания 1а-волокон (Аа-волокна)
Мышечные веретена расположены по всей мышце. Их название обусловлено их веретенообразной формой. Количество мышечных веретен в мышце варьирует от 40 до 500 (Lindel, 2006). Как уже было отмечено, они лежат вдоль мышечных волокон (экстрафузальных волокон, вне веретена) и имеют 5-10 мм в длину и 0,2 мм в толщину (Kolster, 2006). Мышечные веретена состоят из соединительнотканной капсулы, поверхностной слой которой соответствует эндомизию, и 10-20 так называемых интрафузальных мышечных волокон (внутри веретена). Концы веретена прикрепляются к соединительнотканной оболочке мышцы или к сухожилию. Интрафузальные волокна только на концах имеют актиновые и миозиновые филаменты, позволяющие им сокращаться, а в центре волокно способно лишь к растяжению.
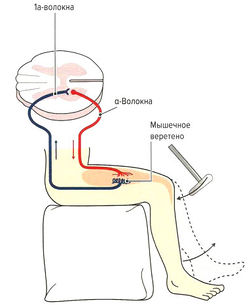
Рис. 1.6 б. Строение и функция мышечного веретена: моносинаптический проприоцептивный рефлекс
Несократительный растяжимый центр интрафузального волокна окружен толстым спиралевидным миелинизированным нервным окончанием типа 1а (Аа-волокна) (кольцеспиральное окончание) (рис. 1.6, а). Диаметр la-волокон составляет 10-20 мкм. Данные нервные волокна являются самыми быстрыми в организме человека — скорость распространения импульса составляет 60-120 м/с. Сократительные концы интрафузальных волокон иннервируются эфферентными у-волокнами (Ay-волокна, у-мотонейроны). Диаметр этих волокон составляет 4-8 мкм, а скорость передачи импульса — 20-50 м/с. Около 50 % всех мотонейронов передних рогов одного сегмента спинного мозга представлены у-мотонейронами (Cabri, 1999).
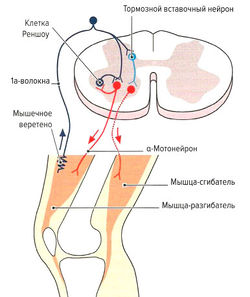
Рис. 1.6 в. Строение и функция мышечного веретена: реципрокное и возвратное ингибирование
Растяжение центральной части интрафузальных волокон приводит к раздражению кольцеспиральных нервных окончаний мышечного веретена (рецептор растяжения). На конце афферентных волокон возникает потенциал действия, который затем передается в ЦНС. Центральная часть веретена растягивается либо при растяжении всей мышцы, либо при напряжении сократительных концов интрафузальных волокон при активации у-мотонейронов. При одновременном действии этих двух факторов возможно как взаимное усиление, так и уменьшение их действия. Таким образом, мышечные веретена способны регистрировать как изменение длины мышцы, так и скорость ее изменения, а также косвенно ее изменять. Действуя через различные контуры регуляции, мышечные веретена выполняют несколько функций.
- Защита мышцы от перерастяжения за счет рефлекса растяжения (моносинаптический рефлекс). Функцией моносинаптического рефлекса растяжения является поддержание мышечной длины. Любое растяжение мышцы, например при ударе по сухожилию или мышце (рис. 1.6, б) или при изменении позы туловища, влияет на натяжение мышечных веретен. В результате резко повышается частота импульсации по 1а-афферентным нервным волокнам, идущим через задние спинномозговые корешки в серое вещество спинного мозга и через один синапс (моносинаптически) связываются с а-мотонейронами этой же мышцы (проприоцептивный рефлекс) (рис. 1.6, б). а-Мотонейроны по а-волокнам вызывают сокращение экстрафузальных волокон натянутой мышцы. Поскольку рецептор и эффектор находятся в одном и том же органе, скорость данного рефлекса очень высока (около 30 мс). Непроизвольные изменения длины мышцы могут сразу корректироваться на уровне спинного мозга без вовлечения высших моторных центров. Одновременно с этим через тормозной вставочный нейрон (ингибирующий la-нейрон) происходит ингибирование мотонейронов мышц-антагонистов (полисинаптический рефлекс) (реципрокное ингибирование антагонистов) (рис. 1.6, в). Благодаря этому растянутая мышца не преодолевает сопротивление антагонистов и предотвращается аналогичная реакция антагонистов при их растяжении (Weiss, 2000). После сокращения растянутой мышцы импульсация от la-волокон снижается. Дополнительно вследствие активации а-мотонейронов через коллатеральные связи и тормозные вставочные нейроны (клетки Реншоу) происходит обратное ингибирование самих а-мотонейронов (рис. 1.6, в). Эта отрицательная связь останавливает распространение сигнала и обеспечивает кратковременность сокращения мышцы, поэтому, несмотря на постоянные внешние раздражители (растяжение мышцы), возможно поддержание ее постоянной длины. Кроме этого, остановка рефлекторного ответа обеспечивается сухожильными органами Гольджи, которые активируются при мышечном сокращении и последующем растяжении сухожилия (аутогенное ингибирование) (см. рис. 1.7, б). В заключение необходимо отметить, что возбудимость мотонейронов контролируется высшими центрами головного мозга. При повреждении этих центров или отходящих от них нервных путей (например, при параплегии) развивается аномальное повышение рефлексов (см. рис. 1.6, а).
- Регуляция активного мышечного тонуса. Высшие моторные центры в головном мозге через так называемые экстрапирамидные пути активируют у-мотонейроны и через мышечные веретена регулируют длину мышц и, следовательно, активный мышечный тонус. Активация у-мотонейронов вызывает напряжение сократительных концов интрафузальных волокон, что растягивает центр мышечных веретен и приводит к активации la-волокон. Как уже было отмечено, импульс затем передается через задние рога в передние рога спинного мозга и возбуждает а-мотонейроны. а-Мотонейроны, в свою очередь, вызывают сокращение волокон скелетных
мышц, что опосредованно уменьшает натяжение центральной части веретен. Эта рефлекторная реакция продолжается до тех пор, пока 1а-волокна не будут регистрировать натяжение (у-петля). При нормальной позе (сидя, стоя или при ходьбе) в большинстве мышц, особенно в работающих против силы тяжести, регистрируется постоянная невысокая активность афферентных волокон мышечных веретен, что позволяет поддерживать определенный активный мышечный тонус. Мышцы, действующие против силы тяжести, например мышца, разгибающая позвоночник, в положении стоя находятся в постоянном небольшом натяжении, т. к. туловище под влиянием силы тяжести наклоняется несколько кпереди. За счет постоянной активации у-мотонейронов достигается дополнительное растяжение средней части мышечных веретен, что вызывает активацию высших моторных центров для поддержания мышечного тонуса. Благодаря постоянной фоновой афферентной импульсации во время обычных движений мышечные веретена могут воспринимать как укорочение, так и растяжение мышцы. При усилении или уменьшении активности у-мотонейронов изменяется чувствительность мышечных веретен и регулируется их активность при конкретных позе и движении. Таким образом, у-мотонейроны устанавливают в каждой конкретной ситуации оптимальную рабочую область мышечных веретен, и они могут регистрировать мышечную длину и скорость ее изменения. Это позволяет устанавливать и поддерживать постоянный активный мышечный тонус, а следовательно, поддерживать равновесие и положение суставов и определенную позу (чувство позы и опоры).
- Выполнение точных движений и регуляция тонкой моторики. Возможность регулировать чувствительность мышечных веретен, особенно вместе с произвольными движениями, играет большую роль. При произвольных движениях, например при разгибании в коленном суставе, происходит помимо активации а-мотонейронов и повышение активности у-мотонейронов. За счет а-у-коактивации устанавливается оптимальная длина мышечных волокон, чтобы поддерживать их чувствительность во время всего движения. При сокращении регистрируется растяжение мышцы и становятся возможными очень точные движения. Крохме того, в научной среде продолжаются дискуссии, являются ли точные движения конечностями исключительно результатом изменения у-активности. Супраспинальные нервные центры могут изменять у-активность в у-петле (изменение частоты импульсации по 1а-волокнам) и таким образом изменять активность связанных с ними а-мотонейронов (Weiss, 2005).
![]() Запомните: Чем больше мышечных веретен в мышце, тем более точные и дифференцированные движения возможны при ее сокращении. Например, в мышцах кисти и глазного яблока находится большое количество проприорецепторов (Kolster, 2005).
Запомните: Чем больше мышечных веретен в мышце, тем более точные и дифференцированные движения возможны при ее сокращении. Например, в мышцах кисти и глазного яблока находится большое количество проприорецепторов (Kolster, 2005).
Сухожильные органы Гольджи[править | править код]
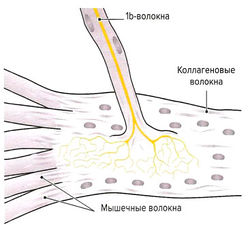
Рис. 1.7 а. Сухожильные органы Гольджи
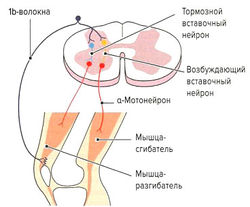
Рис. 1.7 б. Принцип аутогенного торможения
Помимо мышечных веретен, которые регулируют длину мышцы, в поперечнополосатой мышечной ткани находятся сухожильные органы Гольджи, регулирующие напряжение мышцы. Они располагаются вдоль экстрафузальных мышечных волокон в области мышечно-сухожильных соединений (рис. 1.7, а) и вплетаются в коллагеновые волокна сухожилия. Органы Гольджи состоят из множества разветвляющихся терминальных окончаний аксона lb-волокон (Аа-волокна), окруженных общей соединительнотканной капсулой. Эти рецепторы имеют в длину около 100 мкм и прикрепляются к 10-15 сухожильным волокнам (Cabri, 1999). При сокращении мышцы коллагеновые волокна натягиваются и сближаются. Расположенные между коллагеновыми волокнами нервные окончания сдавливаются и генерируют потенциал действия, который в виде нервного импульса передается по lb-волокнам в ЦНС. Адекватньш раздражителем для сухожильных органов Гольджи является изменение натяжения сухожилия, что происходит как при укорочении, так и при растяжении мышцы. Порог чувствительности этих рецепторов выше, чем у мышечных веретен. В отличие от моносинаптической дуги веретен, на уровне спинного мозга переключение импульса на а-мотонейрон идет через два или три синапса. Через тормозные вставочные нейроны (ди- или трисинаптически) происходит ингибирование а-мотонейронов заинтересованной мышцы аутогенное торможение). Одновременно с этим органы Гольджи через два синапса активируют через возбуждающие вставочные нейроны а-мотонейроны мышцы-антагониста (рис. 1.7, б). Оба этих механизма защищают мышцу от избыточного натяжения (защита от перегрузки).
Читайте также[править | править код]
- Поперечно-полосатые мышцы
- Кровоснабжение скелетных мышц
- Иннервация поперечно-полосатых мышц
- Cтроение и функции нейрона
- Синаптическая передача
- Нервно-мышечная передача
- Двигательная единица мыщцы
- Мышечно-сухожильное соединение
- Сухожилие
- Костно-сухожильное соединение
- Типы мышц
- Типы мышечных волокон
- Функции поперечно-полосатых скелетных мышц
- Гладкие мышцы
- Сердце
Источник
Чувствительная иннервация скелетных мышц: нервно-мышечные веретена, сухожильные органы Гольджи, свободные нервные окончания
а) Нервно-мышечные веретена. Разные мышцы содержат от десятков до нескольких сотен мышечных веретен, длина которых не превышает 1 см. Большое количество мышечных веретен расположено в (а) «антигравитационных» (постуральных) мышцах позвоночника, бедра и голени, а также (б) мышцах шеи и (в) внутренних мышцах руки. Все перечисленные мышцы содержат множество медленных оксидативных волокон. Мышечные веретена представлены в небольшом количестве в мышцах, состоящих преимущественно из быстрых гликолитических и промежуточных волокон.
Мышечные веретена состоят из интрафузальных мышечных волокон, количество которых не превышает 12. (В данном контексте экстрафузалъные мышечные волокна считают обычными). Крупные интрафузальные волокна начинаются от полюсов (концов) мышечных веретен и фиксируются к соединительной ткани — перимизию. Некрупные интрафузальные волокна фиксируются к коллагеновой капсуле веретена. В средней части (экваторе) веретена саркомеры практически полностью замещены ядрами, расположенными в форме сумки—сумчато-ядерные мышечные волокна (характерно для широких волокон) или цепочки — цепочечно-ядерные мышечные волокна (характерно для тонких волокон).
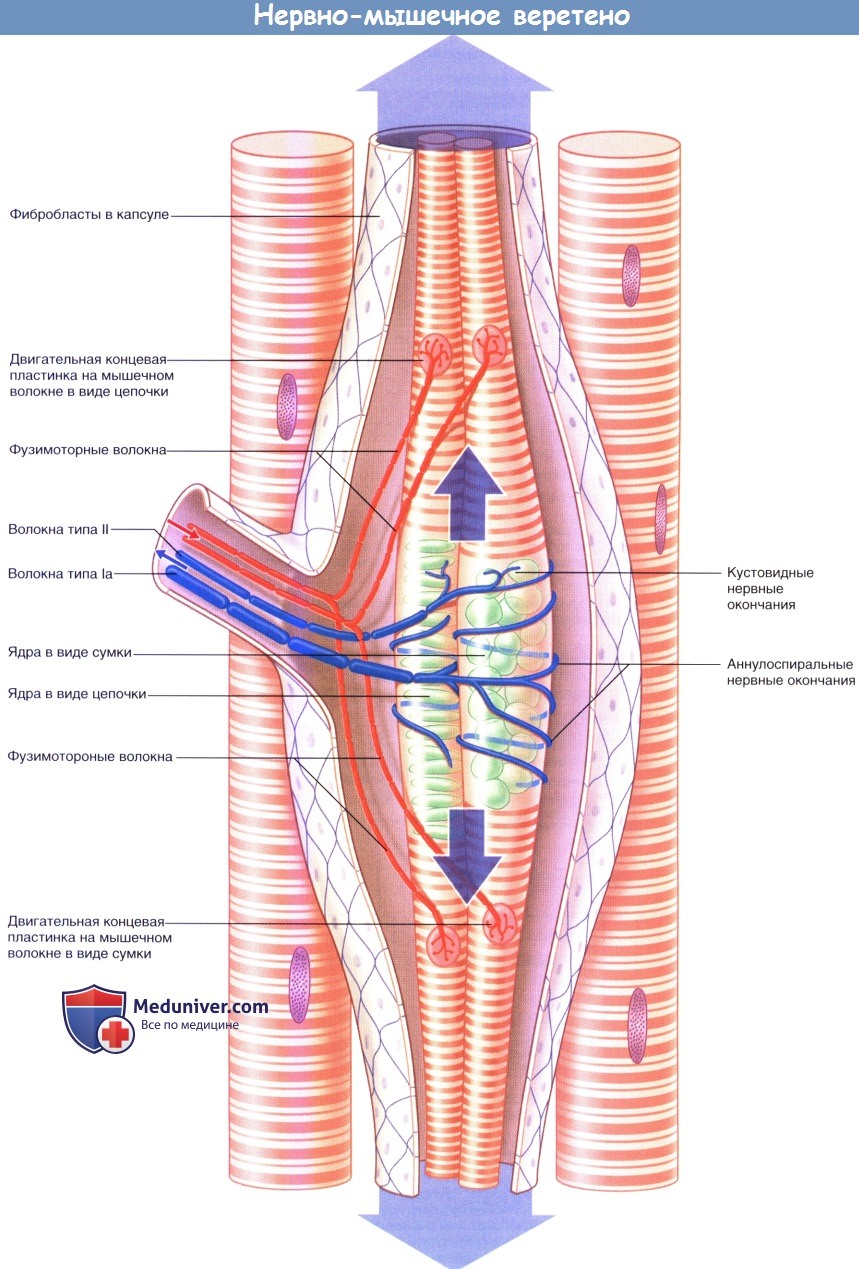
Упрощенное строение нервно-мышечного веретена. Большими стрелками обозначено пассивное растяжение аннулоспиральных нервных окончаний при удлинении мышцы в состоянии покоя.
Стрелками среднего размера обозначено активное растяжение аннулоспиральных нервных окончаний фузимоторными нервными волокнами.
Активное растяжение в достаточной степени компенсирует эффект разгрузки одновременного сокращения экстрафузальных мышечных волокон.
Стрелками маленького размера показаны направления проведения импульсов к мышечному веретену и от него при сокращении мышцы.
1. Иннервация. Иннервацию мышечных веретен осуществляют двигательные и чувствительные нервные волокна. Двигательные нервные волокна — фузимоторные—в соответствии с толщиной относят к типу Аγ, а нервные волокна экстрафузальных мышц — к типу Аα. Фузимоторные аксоны разветвляются и иннервируют поперечно-исчерченные сегменты концевых участков интрафузальных мышц. Единичные первичные чувствительные нервные волокна типа 1а оборачиваются спирально-кольцевым образом вокруг сумчато-ядерных или цепочечно-ядерных мышечных волокон. Вторичные кустовидные чувствительные нервные окончания образованы нервными волокнами II типа и располагаются с одной или с двух сторон от первичного волокна.
2. Активация. Мышечные веретена представляют собой рецепторы растяжения. При растяжении мышечного веретена ионные каналы поверхностной мембраны чувствительных нервных окончаний открываются, за счет чего создаются положительные электроволны. В результате их суммирования в области терминального участка аксона чувствительного нервного волокна происходит образование рецепторного потенциала, который при достижении пороговых значений генерирует нервные импульсы.
Растяжение мышечных веретен может происходить пассивным и активным путями.
3. Пассивное растяжение. Пассивное растяжение мышечного веретена происходит при пассивном удлинении всего мышечного брюшка. Например, при воспроизведении сухожильных рефлексов, в частности коленного, во время удара по сухожилию происходит пассивное растяжение мышечных веретен брюшка четырехглавой мышцы бедра. Волокна типов Iа и II направляются к спинному мозгу и образуют синапсы с дендритами α-мотонейронов. (Принадлежность мотонейронов к типу а определяется Аα-диаметром их аксонов.)
По механизму положительной обратной связи происходит резкое сокращение экстрафузальных волокон четырехглавой мышцы бедра, что приводит к пассивному укорочению мышечного веретена, поскольку оно лежит параллельно экстрафузальным волокнам. Вследствие того, что мышечные веретена лежат параллельно экстрафузальным мышцам, их сокращение происходит пассивно. Этот процесс условно описывают как «разгрузка мышечного веретена».
Сухожильные рефлексы относят к моносинаптическим. Для них характерен латентный период (промежуток времени до ответной реакции на стимул), составляющий 15-25 мс.
Помимо возбуждающего действия на гомонимные мотонейроны (т. е. двигательные нейроны, иннервирующие одни и те же мышцы), афферентные волокна мышечных веретен опосредованно через вставочные тормозные мотонейроны оказывают тормозное действие на а-мотонейроны мышц-антагонистов. Этот процесс получил название реципрокного торможения. Задействованные тормозные нейроны называют вставочными нейронами типа Iа.
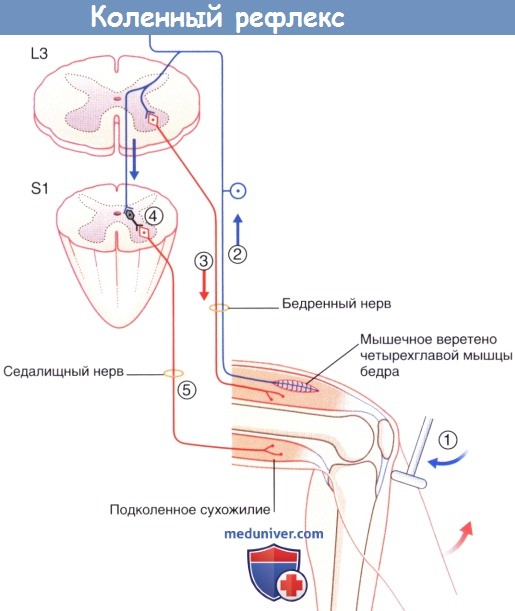
Коленный рефлекс, в котором задействовано реципрокное торможение.
Стрелочками указано проведение нервных импульсов.
(1) Удар по коленной связке вызывает растяжение мышечных веретен в четырёхглавой мышце спины.
(2) Мышечные веретена посылают возбуждающий потенциал в спинной мозг.
(3) α-Мотонейроны вызывают подергивание четырехглавой мышцы и разгибание колена.
(4 и 5) Тормозные вставочные нейроны типа Ia полностью подавляют активность подколенного сухожилия.
4. Кодирование информации. Первичные афференты мышечных веретен проявляют наибольшую активность во время растяжения. Они определяют скорость растяжения мышцы: чем быстрее происходит растяжение, тем больше импульсов они генерируют.
Вторичные афференты мышечных веретен по сравнению с первичными активнее задействованы в процессе поддержания определенной позиции и определяют степень растяжения мышцы: чем выше степень статического растяжения, тем больше формируется импульсов.
5. Активное растяжение. Активное растяжение осуществляют фузимоторные нейроны, вызывающие сокращение поперечно-исчерченных сегментов интрафузальных мышечных волокон. Интрафузальные волокна фиксированы к соединительной ткани и растягивают среднюю часть (экватор) по направлению к полюсам. Это явление получило название «эффект новогодней хлопушки».
В процессе произвольных движений за счет кортикоспинального (пирамидного) проводящего пути происходит одновременная активация Аα- и Аγ-мотонейронов. Таким образом, под действием сокращения экстрафузальных мышц не происходит «разгрузка мышечных веретен». Афференты мышечных веретен, расположенные по обеим сторонам соответствующих суставов, направляют в мозг сигналы обо всех происходящих в данный момент времени сокращениях и расслаблениях мышц.
Активное растяжение мышечного веретена в изометрических условиях.
Термин «изометрический» обозначает «имеющий постоянную длину». Экстрафузальные мышечные волокна находятся в изометрическом состоянии, если они растянуты и их концы зафиксированы.
Мышечное веретено также сохраняет постоянную длину за счет того, что оно опосредованно через соединительную ткань прикрепляется к пластинке.
Однако поперечно-исчерченные компоненты интрафузальных волокон не поддерживаются в изометрическом состоянии: они укорачиваются, поскольку их центральная часть эластична и подвержена внешнему воздействию.
Первичные и вторичные афференты мышечного веретена, подходящие к его центральной части, обеспечивают «активное» растяжение за счет фузимоторной активности, осуществляемой путем направления афферентных импульсов в ЦНС и последующего усиления экстрафузальных сокращений с задействованием механизма гамма-петли.
б) Нервные окончания сухожилий. Сухожильные органы Гольджи располагаются в местах соединения сухожилий с мышцами. Единичное нервное волокно типа Ib распадается на сложноорганизованные мельчайшие разветвления, оплетающие пучки сухожильных волокон, связанные с соединительно-тканной капсулой.
Десятки мышечных волокон погружаются в сухожильные интракапсулярные волокна, которые последовательно связываются с другими мышечными волокнами в пределах определенной мышцы. Под действием натяжения, возникающего в процессе сокращения мышцы, происходит активация луковицеобразных нервных окончаний. В связи с тем, что скорость возникновения импульсов в исходном нервном волокне зависит от силы натяжения, сухожильные нервные окончания определяют силу сокращения мышцы.
Афференты типа Iб передают сигнал на гомонимные мотонейроны по механизму отрицательной обратной связи, в отличие от афферентов мышечных веретен, передающих информацию посредством положительной обратной связи. Это явление получило название «аутогенное торможение»; рефлекторная дуга в данном случае — двухсинаптическая, поскольку в ее образовании принимает участие вставочный тормозной нейрон. При необходимости этот процесс сопровождает реципрокное возбуждение мотонейронов, иннервирующих мышцы-антагонисты.
Важная функция сухожильного органа Гольджи — ограничение и «сглаживание» колебательных движений, характерных для совершающих движение сегментов конечности. Такое ограничение известно с точки зрения физиологии как «скованность в суставах». Парадоксально излишняя активность афферентов типа Iб, типичная для болезни Паркинсона, усиливает колебательные движения, что проявляется характерным тремором покоя, который наиболее выражен в области предплечья (движения пронации и супинации) и пальцев (большой палец совершает движения, напоминающие пересчитывание монет или катание пилюль).
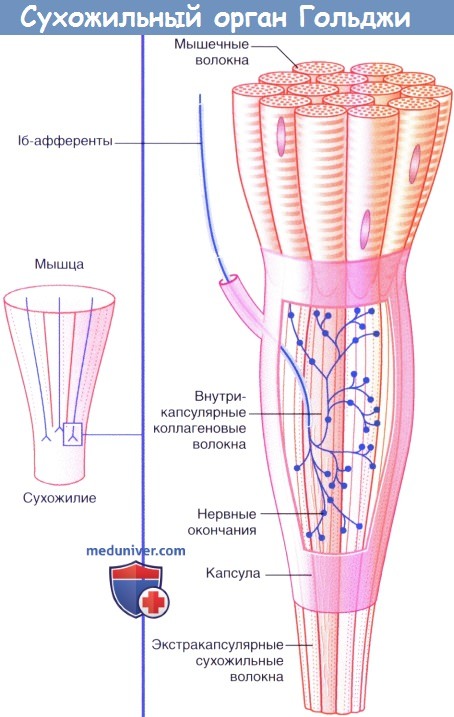
Сухожильный орган Гольджи.
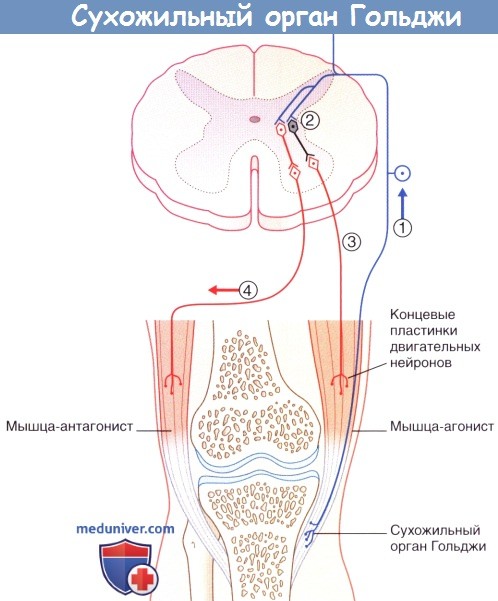
Рефлекторные эффекты при стимуляции сухожильного органа Гольджи.
(1) Сокращение мышцы-агониста вызывает возбуждение афферентов сухожильного органа Гольджи,
что приводит к (2) возбуждению тормозных вставочных нейронов, образующих синапсы с гомонимными мотонейронами,
а также (3) возбуждению тормозных вставочных нейронов, образующих синапсы с (4) мотонейронами мышцы-антагониста.
в) Свободные нервные окончания. В мышцах имеется множество свободных нервных окончаний, большинство которых расположено во внутримышечной соединительной ткани и покрывающих мышцы фасциях. Эти нервные окончания отвечают за болевую чувствительность, возникающую при непосредственном повреждающем воздействии или при накоплении продуктов распада, к которым относят молочную кислоту.
г) Иннервация суставов. Немиелинизированные нервные волокна со свободными нервными окончаниями в большом количестве присутствуют в связках и капсулах суставов, а также во внешних частях внутрисуставных менисков. Эти нервные волокна обеспечивают болевую чувствительность при деформациях суставов, а также участвуют в формировании защитного рефлекса для капсулы сустава. Так, например, переднюю капсулу запястья иннервируют срединный и локтевой нервы; внезапное растяжение при насильственном разгибании вызывает рефлекторную активацию двигательных волокон, что приводит к сгибанию верхней конечности в лучезапястном суставе.
В экспериментах на животных показано, что при воспалении сустава в возбужденном состоянии находится большее количество нервных волокон, чем при растяжении капсулы здорового сустава. Вероятно, существуют нервные окончания, единственным стимулом которых служит воспалительный процесс.
Инкапсулированные нервные окончания, расположенные внутри суставных капсул и вокруг них, представлены тельцами Руффини, реагирующими на натяжение, ламеллярными окончаниями, воспринимающими давление, а также тельцами Пачини, ответственными за ощущение вибрации.
д) Миофасциальный болевой синдром. Миофасциальный болевой синдром—распространенное заболевание, проявляющееся региональной мышечной болезненностью, связанной с чрезмерной чувствительностью пучков напряженных мышечных волокон. (Сходные синдромы наблюдают при другом заболевании—фибромиалгии. Однако при фибромиалгии происходит центральное нарушение болевой чувствительности—дисфункция системы модуляции боли.)
Прикосновение к пучкам мышечных волокон с повышенной чувствительностью вызывает боль; клинически эту область называют миофасциальной триггерной точкой. Для боли не характерно распространение в области дерматома какого-то определенного нерва; в некоторых случаях боль может выходить за пределы триггерной точки—иррадиировать. Кроме того, боль могут сопровождать вегетативные проявления, такие как покраснение и пилоэрекция. Возникновение триггерных точек может быть связано с травмой мышц, чрезмерными нагрузками во время профессиональной деятельности или спортивных занятий при нарушении процесса нормального восстановления.
Спонтанно активирующиеся очаги получили название активных миофасциальных триггерных точек (МТТ), а находящиеся в данный момент времени в неактивном состоянии—латентных миофасциальных триггерных точек. Происходящие процессы недостаточно ясны с точки зрения патофизиологии, однако установлено, что тканевая жидкость, окружающая активные МТТ, содержит большее количество ассоциированных с воспалением веществ (например, брадикинины, простагландины, протоны Н+).
Со временем боль может захватывать новые области или усиливаться в результате раздражения нейронов задних рогов. Высвобождение другими ветвями раздраженных нейронов субстанции Р может привести к возникновению новых МТТ в области той же или прилежащей мышцы.
Стойкое сокращение мышечных волокон, прилежащих к узелковым утолщениям, объясняют инактивацией ацетилхолинэстеразы в базальной мембране их концевых двигательных пластинок. Принципы лечения этого заболевания—длительное пассивное растяжение пораженных мышц, длительное давление в горизонтальном положении больного (например, путем подкладывания теннисного мячика под пораженную область), а также механическое повреждение прокалыванием иглой или введением местных анестетиков и/или стероидов.
е) Резюме. Мышцы. Двигательная единица состоит из двигательного нейрона и иннервируемой им группы мышечных волокон. Каждая двигательная единица содержит определенный гистохимический вид мышечных волокон. Концевое утолщение нервно-мышечного синапса, содержащее пузырьки с АХ, отделено от синаптических складок сарколеммы базальной мембраной, в которой присутствует ацетилхолинэстераза.
В состав мышечных веретен входят интрафузальные мышечные волокна, активация которых происходит с обоих концов за счет γ-фузимоторных нейронов. Чувствительные волокна типа Iа образуют первичные аннулоспиральные нервные окончания в области средней части (экватора), а волокна типа II — вторичные нервные окончания. Оба типа волокон являются рецепторами растяжения. Растяжение может происходить пассивным (например, при сухожильном рефлексе) или активным путем во время фузимоторной активности. Гомонимные мотонейроны—моносинаптические; мышцы-антагонисты реципрокно ингибируются посредством вставочных нейронов Iа. Первичные афференты мышечных веретен определяют скорость мышечного сокращения, а вторичные — степень.
В процессе произвольных движений происходит одновременная активация А α- и Аγ-мотонейронов.
Сухожильные органы Гольджи определяют силу мышечных сокращений. В состав сухожильных органов входит инкапсулированное сухожилие, иннервацию которого осуществляют афференты типа Iб, вызывающие двухсинаптическое торможение гомонимных мотонейронов и реципрокное возбуждение мышц-анта-гонистов.
Свободные внутримышечные нервные окончания обеспечивают болевую чувствительность.
Суставы. Свободные нервные окончания в большом количестве присутствуют в связках и капсулах суставов, а также во внешних частях внутрисуставных менисков. Они обеспечивают болевую чувствительность и формирование суставного защитного рефлекса. Инкапсулированные нервные окончания принимают импульсы от движений суставов.
— Вернуться в оглавление раздела «Неврология.»
Редактор: Искандер Милевски. Дата публикации: 12.11.2018
Источник