Мышечные веретена или рецепторы растяжения

Можно выделить два основных вида специфических рецепторов, расположенных в скелетной мышце: мышечные веретена и рецепторы Гольджи (сухожильные рецепторы Гольджи). В этой статье рассмотрено строение и функции одного вида рецепторов скелетной мышцы – мышечных веретен. Описаны строение и функции интрафузальных мышечных волокон, входящих в состав мышечных веретен.
РЕЦЕПТОРЫ СКЕЛЕТНОЙ МЫШЦЫ (МЫШЕЧНЫЕ ВЕРЕТЕНА)
Познакомившись с строением нервов, можно обсудить вопрос о рецепторах скелетной мышцы. Можно выделить два основных вида рецепторов, расположенных в скелетной мышце: мышечные веретена и рецепторы Гольджи (сухожильные рецепторы Гольджи).
Мышечные веретена были описаны в середине XIX века немецким физиологом Вильгельмом Кюне (Wilhelm Kühne, 1863). Мышечное веретено представляет собой вытянутую структуру, расширенную посередине, что делает его похожим на веретено, которое в давние времена использовали, когда пряли. Английский физиолог, лауреат Нобелевской премии Чарльз Скотт Шеррингтон одним из первых указал, что мышечные веретена являются чувствительными рецепторами мышцы.
Мышечные веретена расположены внутри мышцы параллельно мышечным волокнам и прикрепляются к эндомизию, окружающему мышечные волокна или к сухожилию. Длина мышечного веретена достигает 10 мм. Ширина мышечного веретена в наиболее широкой его части – капсуле колеблется от 80 до 250 мкм (рис.1)
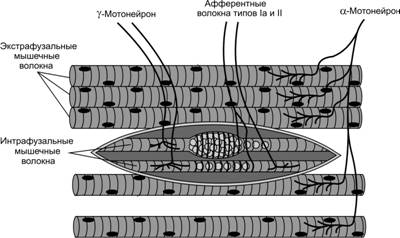
Рис.1. Строение мышечного веретена
Более подробно строение и функции мышц описаны в моих книгах «Гипертрофия скелетных мышц человека» и «Биомеханика мышц»
Количество мышечных веретен в мышце колеблется от нескольких десятков до нескольких сотен. Так, в двуглавой мышце плеча имеется 320 веретен, в большой грудной – 450, а в трехглавой мышце плеча – 520. Плотность веретен (то есть количество веретен приходящихся на 1 г массы мышцы) сильно варьирует. Она наибольшая в тыльных мышцах шеи и наименьшая в мышцах конечностей. Так, например в верхней косой мышце головы плотность веретен составляет 42,7 на 1 г массы мышцы, а в двуглавой мышце плеча – 2,0 на 1 г массы мышцы.
Установлено, что мышечные веретена возбуждаются при растягивании мышцы. Растяжение мышцы возникает, когда, например, занимающиеся выполняют упражнения на растяжение или упражнения в уступающем (эксцентрическом режиме). Также растяжение мышц возникает при локомоциях человека – ходьбе, беге, прыжках или езде на велосипеде.
Информация, поступающая от рецепторов мышечных веретен, сообщает нервной системе о том, какова в настоящий момент времени фактическая длина мышцы (ее еще называют текущей длиной – то есть длиной в текущий момент времени) и скорость растяжения мышцы.
Внутри мышечного веретена располагаются тонкие мышечные волокна, которые называются интрафузальными. Их диаметр в 2-3 раза меньше обычных мышечных волокон, называемых экстрафузальными. Интрафузальные мышечные волокна очень маленькие, однако они также могут сокращаться и расслабляться. Интрафузальных мышечных волокон в мышечном веретене немного: от 4 до 14. Следует отметить, что интрафузальные мышечные волокна в веретене имеют собственную иннервацию, которая позволяет регулировать чувствительность мышечного веретена. К интрафузальным мышечным волокнам подходят двигательные мотонейроны (γ-мотонейроны) и от них отходят чувствительные волокна, несущие информацию в ЦНС о длине и скорости растяжения мышцы. За открытия в этой области шведскому физиологу Рагнару Граниту была присуждена Нобелевская премия.
Благодаря информации, поступающей от мышечных веретен, предотвращается (правда не всегда) повреждение мышцы. Эта информация служит основой для функционирования рефлекса на растяжение.
Литература
- Гранит Р. Основы регуляции движений. – М.: Мир, 1973.– 278 с. Ил.
- Мак-Комас, А.Дж. Скелетные мышцы. – Киев: Олимпийская литература, 2001.– 407 с.
- Самсонова А.В. Моторные и сенсорные компоненты биомеханической структуры физических упражнений: автореф. дис… докт. пед. наук.- СПб, 1998.- 48 с.
- Самсонова, А.В. Биомеханика мышц [Текст]: учебно-методическое пособие /А.В. Самсонова Е.Н. Комиссарова /Под ред. А.В. Самсоновой /Санкт-Петербургский гос. Ун-т физической культуры им. П.Ф. Лесгафта.- СПб,: [б.н.], 2008.– 127 с.
- Самсонова, А.В. Гипертрофия скелетных мышц человека. – СПб: Кинетика, 2018.– 159 с.
С уважением, А.В.Самсонова
Источник
Мышечное веретено. Иннервация мышечного веретена
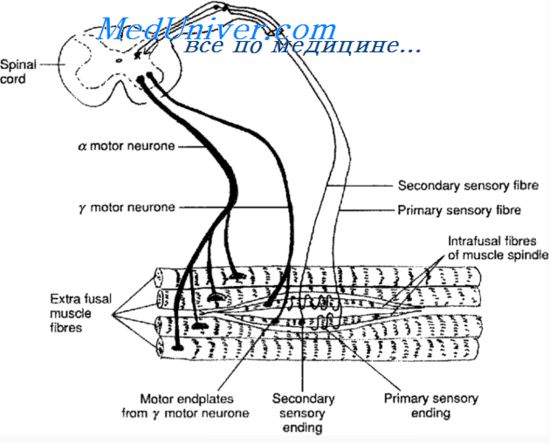
Организация мышечного веретена представлена на рисунке. Каждое веретено имеет длину 3-10 мм и состоит из 3-12 очень маленьких заостренных на концах интрафузалъных мышечных волокон, которые прикрепляются к гликока-ликсу окружающих крупных экстрафузалъных волокон скелетной мышцы.
Интрафузальное мышечное волокно — очень маленькое поперечнополосатое волокно. Однако центральная часть каждого из них, т.е. средняя область между их концами, практически не содержит актиновых и миозиновых нитей. Следовательно, при сокращении концов такого волокна центральная его часть не сокращается. Вместо этого она функционирует как сенсорный рецептор, что обсуждается далее.
Способные сокращаться концевые части интрафузальных волокон возбуждаются тонкими гамма-двигательными нервными волокнами (А), которые являются аксонами небольших гамма-мотонейронов передних рогов спинного мозга, изложенных ранее. Эти волокна называют также гамма-эфферентными волокнами в отличие от крупных альфа-эфферентных волокон (нервные волокна типа А), иннервирующих экстрафузальные волокна скелетных мышц.
Сенсорная иннервация мышечного веретена. Рецепторной частью мышечного веретена является его центральная часть. В этой области интрафузальные мышечные волокна не имеют миозиновых и актиновых сократительных элементов. Они стимулируются растяжением средней части веретена. Легко понять, что рецептор мышечного веретена может возбуждаться двумя путями.
1. Удлинение всей мышцы растягивает среднюю часть веретена и, следовательно, возбуждает рецептор.
2. Даже если длина всей мышцы не изменяется, сокращение конечных частей интрафузальных волокон веретена растягивает его среднюю часть и таким образом возбуждает рецептор.
В центральной рецепторной части мышечного веретена обнаруживают два типа сенсорных окончаний: первичные и вторичные.
Первичное окончание. Центральную часть рецепторной области каждого интрафузального волокна окружает толстое чувствительное нервное волокно, формирующее так называемое первичное, или аннулоспиральное, окончание. Это нервное волокно типа Iа со средним диаметром 17 мкм проводит сенсорные сигналы к спинному мозгу со скоростью 70-120 м/сек — самой высокой скоростью для нервных волокон нашего организма.
Вторичное окончание. Как видно на рисунке, рецепторная область иннервируется обычно одним, но иногда двумя более мелкими сенсорными нервными волокнами типа II со средним диаметром 8 мкм, расположенными с одной или обеих сторон от первичного окончания. Окончание такого волокна называют вторичным; иногда оно окружает интрафузальные волокна так же, как это делают волокна типа Iа, но часто разветвляются подобно кусту.
Внутри мышечного веретена различают два типа интрафузальных волокон: (1) волокна с ядерной сумкой (в соотношении 1 : 3 в каждом веретене), в центральной части рецепторной области которых несколько ядер мышечного волокна собраны в расширенные сумки, как в верхнем волокне на рисунке; (2) волокна с ядерной цепочкой (3 : 9), которые вдвое меньше в диаметре и вдвое короче по сравнению с волокнами с ядерной сумкой, а их ядра распределены в виде цепочки по всей рецепторной области, как в нижнем волокне на рисунке.
Первичное сенсорное нервное окончание (диаметром 17 мкм) возбуждается интрафузальными волокнами и с ядерной сумкой, и с ядерной цепочкой. С другой стороны, вторичное окончание (сенсорное волокно диаметром 8 мкм) обычно возбуждается только волокнами с ядерной цепочкой.
Реакция первичного и вторичного окончаний на длину рецептора — статический ответ. Когда рецепторная часть мышечного веретена растягивается медленно, количество импульсов, передаваемых от первичных и вторичных окончаний, возрастает прямо пропорционально степени растяжения, и окончания продолжают передавать эти импульсы в течение нескольких минут. Этот эффект называют статическим ответом мышечного веретена. Это значит, что и первичные, и вторичные окончания продолжают передавать сигналы в течение нескольких минут, если само мышечное веретено остается растянутым.
— Вернуться в оглавление раздела «Физиология человека.»
Оглавление темы «Обоняние. Функции спинного мозга»:
1. Вкусовой порог. Вкусовая почка языка
2. Механизм стимуляции вкусовых почек. Нервные пути вкусовой чувствительности
3. Вкусовые предпочтения. Обоняние
4. Стимуляция обонятельных клеток. Адаптация обонятельных клеток
5. Первые обонятельные ощущения. Передача обонятельных сигналов
6. Обонятельный тракт и пути. Новый обонятельный путь
7. Двигательные функции спинного мозга. Спинальные и децеребрированные животные
8. Передние мотонейроны спинного мозга. Вставочные нейроны спинного мозга
9. Тормозная система клеток Реншоу. Мышечные сенсорные рецепторы
10. Мышечное веретено. Иннервация мышечного веретена
Источник
В активной части опорно двигательного аппарата имеются проприорецепторы. Существует два типа проприорецепторов – нервно-мышечные веретёна, которые реагируют на изменение длины мышцы, а также сухожильные органы Гольджи, реагирующие на изменение напряжения, то есть на силу сокращения мышечных волокон (Рисунок 1).

Рисунок 1. Проприорецепторы: нервно-мышечное веретено и сухожильный орган Гольджи
Нервно-мышечные веретёна. Нервно-мышечные веретена расположены в толще скелетных мышц между мышечными волокнами. Они представляют собой веретеновидные структуры, покрытые соединительнотканной капсулой. Длина веретена составляет 4 – 7 (до 10) мм, диаметр – 100 мкм. Под капсулой внутри веретена находятся видоизменённые мышечные волокна, которые называются интрафузальными. Эти волокна имеют ряд отличий от обычных, экстрафузальных, мышечных волокон. Во-первых, интрафузальное волокно имеет меньший диаметр (15 – 30 мкм) по сравнению с экстрафузальным (50 – 100 мкм). Во-вторых, в центральной («экваториальной») части интрафузального волокна нет сократительных элементов, поэтому к сокращению способны только периферические части. Поскольку концы интрафузального волокна фиксированы к соединительнотканной капсуле, покрывающей веретено, при укорочении периферических отделов волокна его центральная часть растягивается. Интрафузальные волокна расположены параллельно эктсрафузальным. По строению интрафузальные волокна делятся на два типа – с ядерной сумкой и с ядерной цепочкой (Рисунок 2). В свою очередь, волокна с ядерной сумкой по функции по особенностям иннервации подразделяются на статические и динамические. Для волокон с ядерной сумкой характерно наличие расширенной центральной части, в которой ядра собраны в скопления. Волокна с ядерной цепочкой примерно вдвое тоньше волокон с ядерной сумкой, ядра в их центральной части расположены в ряд. В каждом веретене содержится 1 – 3 волокна с ядерной сумкой и 5 – 7 волокон с ядерной цепочкой.

Интрафузальные волокна получают как афферентную, так и эфферентную иннервацию. Выделяют два типа афферентных окончаний – первичные и вторичные. Первичные окончания называют аннулоспиральными, они представляют собой спираль, окружающую центральную часть всех интрафузальных волокон. От первичных окончаний отходят волокна типа Ia, которые соответствуют типу Aα по Гассеру-Эрлангеру. Это самые толстые волокна (~17 мкм) с самой высокой скоростью проведения возбуждения (70 – 12 м/с). Каждое нервно-мышечное веретено иннервируется только одним волокном Ia. Другими словами одно афферентное нервное волокно Ia получает импульсацию со всех первичных окончаний в иннервируемом им веретене. Вторичные окончания имеют форму спирали, но могут также образовывать разветвлённую структуру. Они оплетают центральную часть интрафузальных волокон с ядерной цепочкой, а также статических волокон с ядерной сумкой, но их не бывает в динамических волокнах с ядерной сумкой. От вторичных окончаний отходят волокна типа II, соответствующие типу Аβ по Гассеру-Эрлангеру; диаметр этих волокон составляет ~8 мкм, они проводят возбуждение со скоростью 40 – 70 м/с (Рисунок 3).

Рисунок 3. Схема нервно-мышечного веретена
Как первичные, так и вторичные нервные окончания чувствительны к изменению длины интрафузальных волокон, однако характер ответной реакции различается. При медленном растяжении центральной части интрафузальных волокон импульсация появляется как в первичных, так и во вторичных рецепторах. Частота импульсов пропорциональная степени растяжения волокна. Такой ответ афферентных окончаний называют статическим. Однако при быстром внезапном растяжении волокна в первичных (но не вторичных) окончаниях отмечается мощный всплеск импульсации – динамический ответ. Следовательно, волокна Ia передают информацию как об абсолютной длине интрафузальных волокон (то есть о степени растяжения), так о скорости её изменения, тогда как волокна II – только об абсолютной длине. Следует понимать, что растяжение центральной части интрафузальных волокон может быть связано как с удлинением всей мышцы (интрафузальные волокна расположены параллельно экстарфузальным), так и при сокращении периферических отделов волокна (Рисунок 3).

Рисунок 4. Статический и динамический ответы рецепторов нервно-мышечных веретён
Эфферентная иннервация интрафузальных волокон представлена нервными волокнами типа Aγ, которые являются аксонами γ-мотонейронов спинного мозга. Диаметр этих волокон составляет ~5 мкм, скорость проведения возбуждения в них – 15 – 40 м/с. Окончания Aγ волокон подходят к периферическим отделам интрафузальных волокон, при возбуждении γ-мотонейронов концевые участки интрафузальных волокон сокращаются, в результате чего их центральные отделы растягиваются. Выделяют γ-динамические и γстатические волокна, γ-динамические волокна иннервируют динамичекие волокна с ядерной сумкой, их активация повышает динамическую чувствительность окончаний Iа типа.
γ-статические волокна иннервируют волокна с ядерной цепочкой и статические волокна с ядерной сумкой, их возбуждение повышает уровень активности как первичных, так и вторичных окончаний (см. ниже).
Миотатический рефлекс (рефлекс растяжения)
В ответ на увеличение длины мышцы рецепторы нервно-мышечных веретён возбуждаются, и импульсация по волокнам Ia (и при быстром, и при медленном растяжении) или II (при медленном растяжении) поступает в спинной мозг. Войдя в серое вещество спинного мозга, волокно афферентного нейрона разветвляется и образует возбуждающие синапсы практически со всеми α-моторными нейронами, иннервирующими гомонимную (то есть эту же) мышцу. Возбуждение α-моторных нейронов вызывает сокращение экстрафузальных волокон, и мышца укорачивается. Следует отметить, что коллатерали афферентных нейронов переключаются также на вставочные тормозные нейроны, которые угнетают α-мотонейроны мышц антагонистов. Таким образом, растяжение мышцы вызывает её рефлекторное укорочение, при этом мышцы антагонисты расслабляются. Описанный рефлекс с рецепторов нервно-мышечных веретён называется миотатическим, или рефлексом на растяжение (Рисунок 4). Миотатический рефлекс является моносинаптическим, поэтому имеет очень небольшой латентный период (~ 0,9 мс).

Рисунок 5. Миотатический рефлекс с нервно-мышечных веретён
Различают фазический (динамический) и тонический миотатический рефлекс. Фазический рефлекс запускается с первичных нервных окончаний при быстром растяжении мышцы, например, при ударе по сухожилию неврологическим молоточком или при появлении внезапного препятствия совершаемому движению. Тонический рефлекс запускается с вторичных нервных окончаний при медленном (или постоянном) растяжении мышечных волокон; он имеет значение для поддержания вертикальной позы. Когда тело отклоняется от правильного положения, мышцы, участвующие в поддержании позы, растягиваются. В результате возбуждаются как первичные, так и вторичные окончания нервно-мышечных веретён, запускается миотатический рефлекс, и мышцы сокращаются; за счёт их сокращения тело возвращается в правильное положение. Когда у длительно стоящего человека (например, солдата в стойке «смирно») из-за утомления сгибаются ноги в коленных суставах, проприорецепторы четырёхглавых мышц реагируют на растяжение, и ноги снова выпрямляются. Постоянное стимулирующее влияние на α-мотонейроны со стороны афферентных волокон Ia и II типов вносит вклад в поддержание мышечного тонуса.
Функции γ-моторных нейронов. Понятие о γ-петле.
Как уже упоминалось, интрафузальные волокна получают эфферентную иннервацию со стороны γ-моторных нейронов. При возбуждении γ-мотонейронов происходит сокращение периферических (концевых) отделов интрафузальных волокон, в результате их центральная часть растягивается. При этом первичные и вторичные нервные окончания активируются, импульсация от них поступает в спинной мозг, и возбуждаются α-моторные нейроны. Другими словами, запускается миотатический рефлекс. Такой механизм (активация рефлекса растяжения при возбуждении γ-мотонейронов) называется γ-петлёй. Наличие γ-петли обеспечивает сохранение чувствительности рецепторов нервномышечных веретён при разной длине мышцы. Во время сокращения мышцы её длина уменьшается, соответственно должны укорачиваться и интрафузальные волокна. В результате импульсация с рецепторов нервно-мышечных веретён должна была бы прекратиться, и рецепторы потеряли бы способность реагировать на изменение длины мышцы. Напротив, при расслаблении мышцы её длина увеличивается, и интрафузальные волокна могут быть чрезмерно растянуты. В таких условиях рецепторы будут генерировать импульсы с максимальной частотой и, следовательно, также потеряют способность реагировать на изменение длины. Однако в реальных условиях имеет место α-γ-коактивация, то есть α- и γмотонейроны возбуждаются одновременно. В результате сокращаются как экстра-, так и интрафузальные волокна. Сокращение экстрафузальных волокон приводит к укорочению мышцы, а сокращение периферических отделов интрафузальных волокон – к растяжению их центральной части. Следовательно, длина центральной части не меняется, поскольку её укорочение вследствие сокращения экстрафузальных волокон компенсируется растяжением из-за сокращения периферических отделов интрафузальных волокон. И наоборот – расслабление всей мышцы сопровождается расслаблением и интрафузальных волокон, при этом растяжение центральной части уменьшается. Таким образом, за счёт α-γ-коактивации рецепторы нервно-мышечных веретён сохраняют чувствительность (и, следовательно, передают информацию о длине мышцы в ЦНС) как при сокращении мышцы, так и при её расслаблении (Рисунок 6). На γ-мотонейронах заканчиваются нисходящие пути из различных отделов головного мозга. Меняя активность γ-мотонейронов, мозговые центры влияют на чувствительность рецепторов нервно-мышечных веретён и, следовательно, на порог рефлекса растяжения. Сухожильные органы Гольджи. Обратный миотатический рефлекс Кроме нервно-мышечных веретён в активной части опорно-двигательного аппарата имеется ещё один тип проприрецепторов – сухожильные органы Гольджи. Сухожильные органы представляют собой сеть коллагеновых нитей, окружённую тонкой соединительнотканной капсулой. Диаметр сухожильного органа Гольджи составляет 100 мкм, длина – 1 мм. Коллагеновые нити внутри органа оплетённых нервными окончаниями, от которых отходят нервные волокна типа Ib. эти нервные волокна, как и Ia, соответствуют типу Aα по Гассеру-Эрлангеру. Сухожильные органы Гольджи расположены в месте прикрепления мышечных волокон к сухожильным, при этом каждый сухожильный орган связан с 3 – 25 (в среднем – с 10 – 15) мышечными волокнами (Рисунок 7). Таким образом, в отличии от нервно-мышечных веретён, которые ориентированы параллельно мышечным
волокнам, сухожильные органы по отношению к мышечным волокнам расположены последовательно.

Рисунок 6. Функция γ-петли
Следовательно, сухожильные органы реагируют не на изменение длины мышцы, а на развиваемое ей усилие, другими словами, на степень её напряжения. Афферентные нервные волокна от сухожильных органов Гольджи следуют в спинной мозг, где, в отличие от чувствительных волокон с нервно-мышечных веретён, переключаются на вставочные тормозные нейроны. Вставочные нейроны, в свою очередь, образуют синаптические контакты с α-мотонейронами гомонимной мышцы. Таким образом, активация сухожильных органов приводит к расслаблению мышцы, к которой крепится данное сухожилие. Описанный рефлекс называется обратным рефлексом растяжения (обратным миотатическим рефлексом) (Рисунок 8). Этот рефлекс, по-видимому, имеет две функции. Во-первых, он предупреждает чрезмерное напряжение скелетных мышц, которое может привести к тяжёлым травмам (разрыву мышцы, отрыву сухожилия от кости или от мышцы). Во-вторых, обратный миотатический рефлекс обеспечивает равномерность силы сокращения разных мышечных волокон в пределах одной мышцы. Те волокна, которые напряжены слишком сильно, расслабляются за счёт усиления тормозных влияний со стороны спинного мозга. Напряжение волокон, которые сокращаются недостаточно сильно, напротив, усиливается за счёт уменьшения тормозного воздействия.

Рисунок 7. Сухожильный орган Гольджи
Проприорецепторы посылают в головной мозг информацию о состоянии скелетно-мышечной системы Импульсация с проприорецепторов поступает не только в спинной мозг, но и по коллатералям афферентных волокон в различные отделы головного мозга. В частности, импульсы с нервно-мышечных веретён и сухожильных органов следуют по спинномозжечковым трактам в мозжечок, а также по системе задних столбов (тонкий и клиновидный пучок) в продолговатый мозг, затем – в таламус, а оттуда – в кору больших полушарий. Таким образом, мозжечок и кора большого мозга постоянно получают информацию о состоянии скелетно-мышечной системы, а именно о длине мышц и о степени их напряжения. Эта информация является необходимой как для регуляции произвольных движений и позы, так и для осознания положения частей тела в пространстве и друг относительно друга.
Рисунок 8. Обратный миотатический рефлекс с сухожильных органов Гольджи
Источник