Рост клетки путем увеличения вакуоли и растяжение клеточной стенки
.png "Рост клетки путем увеличения вакуоли и растяжение клеточной стенки")
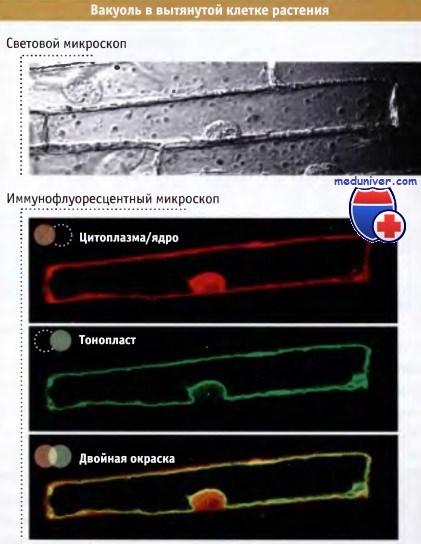
Механизм роста клеток растений набуханием вакуолей• Наполнение вакуолей водой представляет собой уникальный механизм, обеспечивающий увеличение объема клеток за счет изменения давления • Существует более одного типа вакуолей До сих пор мы исключительно рассматривали процессы митоза и цитокинеза, происходящие в апикальной зоне клеточного деления. Что же происходит с клеткой после окончания деления? К числу наиболее очевидных изменений относится увеличение размера клетки, в котором основную роль играют вакуоли. В зоне деления находятся относительно небольшие клетки, содержащие максимум несколько маленьких вакуолей, причем в некоторых клетках они вообще отсутствуют. После выхода клеток из зоны деления они вступают в зону растяжения. Там в результате поступления воды в центральные вакуоли клетки увеличиваются в размере. Этот процесс включает три скоординированных этапа: В статьях на сайте, прежде чем рассматривать участие клеточной стенки в процессе увеличения размера клетки, мы коснемся роли вакуолей в этом процессе. Способность вакуолей увеличивать величину клеток зависит от одной из наиболее существенных физических характеристик последних. Жизнедеятельность клеток зависит от наличия барьера, отделяющего их внутреннее содержимое от окружающей среды. Это дает им возможность обосабливать метаболические реакции таким образом, что они происходят в стабильных условиях, которые не нарушаются изменениями, происходящими в окружающей среде. Клетки растений и животных окружены мембранами, богатыми липидами. При этом макромолекулы и заряженные молекулы через эти мембраны свободно не проходят и, таким образом, имеют возможность концентрироваться внутри клетки. Однако мембраны проницаемы для воды и небольших нейтральных молекул, которые проходят через них благодаря осмосу. Осмос представляет собой процесс, за счет которого через полупроницаемую, подобно клеточной, мембрану проходит вода. Это обеспечивается различием в концентрациях растворенных веществ по обе стороны мембраны. При этом ток воды направляется от менее концентрированного к более концентрированному раствору. Захватывая из окружающей среды соли, а также продуцируя другие молекулы, клетки устанавливают осмотический дисбаланс между цитоплазмой и окружающей средой. При этом через мембраны возникает ток воды, необходимый для уравнивания давления. Из-за притока воды, необходимого для разбавления концентрированного раствора, клетка пребывает в состоянии постоянной опасности «погибнуть от наводнения» и, в конце концов, может лопнуть. Клетки животных постоянно расходуют энергию для откачивания таких ионов, как натрий. Это снижает концентрацию растворенных веществ и противодействует поступлению в клетку воды. Клетки растений для решения этой же проблемы выработали совершенно другую стратегию и используют поток воды для управления своим ростом. У растений поступившая в клетку вода запасается в вакуолях. Активно накапливая из окружающей среды минеральные соли, вакуоли способствуют дальнейшему притоку в клетку воды за счет осмоса, в результате чего сильно разбухают. После того как клетка прекращает деление и расширяется, большая центральная вакуоль может занимать до 95% ее объема, как это, например, показано на рисунке ниже. Поэтому вакуоль вполне можно считать самой крупной органеллой растительной клетки. Окруженная собственной мембраной (тонопластом), центральная увеличенная вакуоль сжимает цитоплазму до размеров тонкого слоя, примыкающего к плазматической мембране. Поскольку вакуоли заполнены, главным образом, водой и минеральными солями, вакуолизация представляет собой экономичный энергосберегающий путь увеличения величины клетки. Хотя вакуоли необходимы для увеличения размеров клеток, они не только просто занимают пространство, но также служат и другим целям. Часто они используются для сохранения небольших молекул, которые или понадобятся клетке позже, или нуждаются в постоянной изоляции от цитоплазмы. Некоторые токсические вещества, попадающие в клетку из окружающей среды, разрушаются или хранятся в вакуолях, которые тем самым выполняют детоксицирующую роль. Иногда растения сами продуцируют токсины в качестве средств защиты от микроорганизмов, и такие токсины запасаются в вакуолях. Некоторые пигменты, окрашивающие различные части растения, также откладываются в вакуолях. Однако не все вещества, попадающие в вакуоли, могут находиться там в течение долгого времени. Вакуоли выполняют роль временных депо для таких необходимых метаболитов, как неорганические ионы и аминокислоты. Существуют вакуоли, которые используются в качестве подобия реакторов, для осуществления метаболических процессов, которые не могут протекать в цитоплазме. Например, литические и пищеварительные вакуоли аналогичны лизосомам клеток животных и вакуолям дрожжей и превращают различные вещества в компоненты, которые усваиваются клеткой. В семенах, вакуоли, в которых хранятся и перевариваются метаболиты, вместе выполняют одну задачу. У них один тип вакуолей выполняет функции накопления и хранения белков, специфических для растений. При созревании в такие вакуоли очень плотно упаковывается один или несколько типов специальных белков. Когда происходит прорастание семени, слияние вакуолей, хранящих белок, и вакуолей, осуществляющих его лизис, приводит к расщеплению белка и к высвобождению аминокислот, которые используются для синтеза белков на ранних стадиях роста растения.
— Вернуться в содержание раздела «генетика» на нашем сайте Оглавление темы «Цитология растений»:
|
Источник
| Ололо Ололоша |

Основой роста тканей, органов и всего растения являются образование и рост клеток меристематической ткани. Различают апикальную, латеральную и интеркалярную (вставочную) меристемы. Апикальная меристема — верхушечная образовательная ткань стеблей и корней. Латеральная меристема (камбий, феллоген) расположена параллельно боковой поверхности органа. Интеркалярная меристема расположена в междоузлиях стебля и в основании листьев растений злаков.
1. Лаг-фаза роста, эмбриональная фаза развития — период между делениями (интерфаза) и собственно деление клетки (митоз). В этот период клетка имеет густую цитоплазму, очень мелкие (зачаточные) вакуоли, много рибосом, мелких митохондрий, пропластид, которые делятся. Первичная клеточная стенка тонкая. В клетках активно идет синтез нуклеиновых кислот, белков, ферментов. В эмбриональных клетках содержание цитокининов (ЦК) больше, чем ауксинов (ИУК).
2. Лог-фаза (рост растяжением, фаза кислого роста). После завершения деления (индолилуксусная кислота). В этой фазе объем клеток быстро увеличивается – в 50—100 и более раз. Мелкие вакуоли и пузырьки аппарата Гольджи сливаются, образуя центральную вакуоль. В вакуоли идет активное, с затратами АТФ, накопление осмотически активных веществ — аминокислот, сахаров, ионов, что приводит к резкому повышению осмотического давления клеточного сока, т.е., к снижению водного потенциала клеток. В вакуоль осмотическим путем поступает вода, объем вакуоли при этом значительно увеличивается, возрастает тургорное давление протопласта на клеточную стенку (КС). Растущие клетки начинают активно синтезировать ИУК, которая индуцирует активный транспорт (выход) ионов Н+ из цитоплазмы в апопласт. Закисление зоны апопласта приводит к разрыхлению клеточной стенки, увеличивает ее пластичность. Одновременно с этим процессом из протопласта путем экзоцитоза поступают фрагменты микрофибрилл целлюлозы, гемицеллюлоз, пектиновых в-в, структурных белков экстенсинов, из которых формируется вторичная многослойная клеточная стенка. В конце фазы растяжения происходит лигнификация клеточных стенок, повышается содержание фенольных ингибиторов и абсцизовой кислоты, снижается содержание ауксина За счет роста клеток растяжением идет увеличение площади листьев, длины стебля и корневой системы растений.
Таким образом, рост клетки растяжением включает:
- разрыхление клеточной стенки;
- осмотическое поступление воды в центральную вакуоль, и увеличение ее объема;
- закрепление увеличения объема путем формирования многослойной вторичной клеточной стенки.
3. Фаза замедленного роста (дифференцировки, этап зрелости). На этом этапе развития клетки приобретают определенные специфические признаки в связи с выполняемыми в дальнейшем функциями (паренхимные, хлорофиллоносные, проводящие, механические, покровные, генеративные клетки). В конце этой фазы окончательно формируется вторичная клеточная стенка у специализированных клеток. В клетке снижается содержание ИУК, возрастает содержание АБК и фенольных соединений.
4. Фаза прекращения роста (этап старения) — завершает онтогенез клетки. В клетках снижается содержание РНК, белков, снижается скорость биосинтетических процессов, возрастает активность ферментов гидролаз, пероксидаз, что ведет к деградации клеточных структур. Возможные причины, вызывающие старение клеток:
- накапливаются повреждения в геноме, в мембранах; в клетках увеличивается концентрация ядовитых веществ.
- Включается генетическая программа старения (PCD — programmed cell death), как последнего этапа онтогенеза.
27.09.2014; 23:18 хиты: 5226 рейтинг:+1
|
Источник
УВЕЛИЧЕНИЕ РАЗМЕРОВ КЛЕТКИ (РОСТ В ФАЗЕ РАСТЯЖЕНИЯ) [c.505]
Механизмы, обусловливающие переход клеток к растяжению, исследованы мало. Предполагается, что большую роль может играть изменение содержания и соотношения фитогормонов и их рецепторов по мере удаления делящейся клетки от инициальных. Выше отмечалось, что для подготовки и перехода клетки к делению необходимо взаимодействие ауксина и цитокинина. Возможно, переход к растяжению вызывается изменением соотношения концентраций ауксина и цитокинина и их рецепторов в сторону, менее благоприятную для деления и более соответствующую росту растяжением, поскольку для роста растяжением обязательно присутствие ауксина. Таким образом, в клетках меристемы, готовящихся перейти к росту растяжением, создаются механизмы, обеспечивающие процесс растяжения. [c.330]
Еще до начала разделения материнской клетки на клеточную пластинку начинают откладываться микрофибриллы целлюлозы. При цитокинезе каждый протопласт выделяет новую первичную оболочку, окружая себя ею. По окончании цикла деления клетка переходит в состояние, называемое рост растяжением . [c.28]
Биогенез митохондрии. Собственная генетическая система и способность митохондриальной ДНК к репликации позволяют митохондриям размножаться самостоятельно. Поэтому в клетке митохондрии образуются из предшествующих митохондрий и, возможно, из промитохондрий. В меристематических клетках обнаруживаются митохондрии, разделенные перетяжкой. При переходе меристематической клетки к росту растяжением число митохондрий в клетке возрастает в 3 — 8 раз и меняется их структура. [c.323]
Влияние ауксинов на рост клетки достаточно хорошо изучено. Рост растяжением предполагает, что целлюлозный каркас клеточной стенки еще не приобрел окончательной жесткости. Увеличение размеров клетки осуществляется как за счет осмотического набухания (растягивания протопласта поступающей в него водой), так и за счет отложения нового материала стенки. Направление растяжения определяется, повидимому, ориентацией уже включенных в нее целлюлозных микрофибрилл. Мягкость стенки поддерживается кислотной средой и ауксинами. В 1973 г. четыре группы ученых независимо друг от друга продемонстрировали, что ауксины стимулируют секрецию протонов из цитоплазмы в клеточную стенку. Это приводит к снижению в ней pH (повышению кислотности) и, следовательно, к ее размягчению . Возможно, некий фермент с низким оптимумом pH разрывает связи в образующих стенку полисахаридах и тем самым обеспечивает растяжимость каркаса в целом. Кроме того, необходимо, чтобы в клетке поддерживался низкий осмотический потенциал и чтобы было достаточно воды, которая проникала бы в клетку и создавала высокое тургорное давление. Согласно более поздним данным, первичное действие ауксинов обусловлено, вероятно, не подкислением клеточной стенки не исключено, что они связываются с рецепторами в плазмалемме эпидермальных клеток, что приводит к изменениям генной активности. Эти изменения приводят к синтезу новых ферментов или других белков, имеющих непосредственное отношение к росту. [c.255]
Несмотря на непрерывное деление клеток, общее их количество в меристеме почти не изменяется. Это связано с тем, что в морфологически нижней части конуса нарастания эмбриональные клетки переходят в следующую фазу роста — растяжение. На второй фазе роста возникают вакуоли. В вакуолярном соке сосредоточиваются разнообразные продукты обмена, возникающие в процессе жизнедеятельности клетки. Наряду с этим в фазе растяжения продолжает существенно увеличиваться и количество протоплазмы. [c.510]
До сих пор обсуждение развития высших растений велось главным образом на уровне описания тканей и органов. Какие же изменения, происходящие на клеточном уровне, лежат в основе всех этих процессов Поскольку клетки растений лишены подвижности из-за наличия клеточных стенок, морфогенез растений должен зависеть от регулируемого деления клеток, сопряженного с ростом клеток в строго определенном направлении. Папример, большинство клеток, образуемых апикальной меристемой корня, проходит три основные фазы развития деление, рост (растяжение) и дифференцировку. Эти фи стадии, во времени и в пространстве накладывающиеся друг на друга, определяют характерное строение кончика корня. Хотя дифференцировка клетки часто начинается, когда она еще увеличивается в размерах, в кончике корня относительно несложно отличить зону деления клеток, зону их растяжения (в результате чего происходит рост корня в длину) и зону дифференцировки (рис. 20-61). После завершения дифференцировки некоторые из дифференцированных типов клеток остаются живыми (например, клетки флоэмы), а другие погибают (например, клетки ксилемы) [c.430]
В цикле выращивания каллусные клетки после ряда делений проходят обычный для клетки растения онтогенез, они приступают к росту растяжением, затем дифференцируются как зрелые каллусные клетки и наконец деградируют. На электронно-микроскопических фотографиях (рис. 4) показана тонкая структура [c.20]
Увеличение размера делящихся клеток происходит за счет Растяжение клеток синтеза структур цитоплазмы и поддерживается поступлением в клетки значительных количеств соединений азота и других питательных веществ. Прекратившие деление клетки многоклеточных растений переходят к более быстрому типу роста — росту растяжением, существующему только у растительных клеток. Он служит важнейшим механизмом, обеспечивающим увеличение площади листовой поверхности, длины стебля и корневой системы, что необходимо для оптимизации процессов питания и других целей. При этом типе роста значительное увеличение объема клеток достигается за счет образования большой центральной вакуоли (см. рис. 1.1). Поглощение воды вакуолью обеспечивается высокой концентрацией осмотически активных веществ в вакуолярном соке. Одновременно с возрастанием объема вакуоли клеточные стенки размягчаются и растягиваются. [c.329]
В конце роста растяжением усиливается лигнификация клеточных стенок, накапливаются ингибиторы фенольной природы и абсцизовая кислота, возрастает активность пероксидазы и оксидазы ИУК, снижающая общее содержание ауксина в тканях. Начавшееся формирование вторичной клеточной стенки лишает клетку способности к растяжению. Все эти факторы приводят к торможению и прекращению удлинения клеток. [c.331]
Рост растяжением включает в себя образование в клетке центральной вакуоли, накопление в ней осмотически активных веществ, поглощение воды и размягчение и растяжение клеточных стенок. Возникающее тургорное давление служит основной силой, растягивающей клеточную стенку. Растяжение клеточных стенок поддерживается включением в них новых молекул полисахаридов. Стенка оказывает на содержимое клетки противодавление, равное тургорному давлению, что определяет тонус клетки и ткани. Одновременно с растяжением клетки синтезируются компоненты цитоплазмы. [c.395]
Растяжение регулируется гормональной системой. Основную роль в регуляции роста растяжением выполняет ИУК. Ауксин вырабатывается в верхушке побега и, перемещаясь в зону растяжения, индуцирует рост клеток, готовых перейти к растяжению. В растягивающихся клетках ИУК взаимодействует с рецепторами, локализованными в цитоплазматической мембране. Одним из результатов этого взаимодействия являются активация выделения ионов Н+ из клеток и подкис-ление фазы клеточных стенок. Одновременно развивается гиперполяризация мембранного потенциала. Таким образом, активация ауксином Н+-ПОМПЫ плазмалеммы является одним из ранних событий в индукции роста растяжением. Подкисление фазы клеточных стенок создает условия для их разрыхления вытеснение Са + из пектатов стенок, ослабление части водородных связей, создание pH, благоприятного для деятельности в стенках кислых гидролаз, увеличивают растяжимость стенок. Одновременно с этим ИУК в комплексе с рецепторами активирует синтез белков и РНК. Активация синтеза РНК и белков необходима для поддержки начавшегося роста растяжением. Усиление дыхания обеспечивает энергетические затраты на все эти процессы. [c.395]
Эта процедура необходима, чтобы гарантировать в вырезанном отрезке колеоптиля только рост растяжением и удалить делящиеся клетки верхушки колеоптиля. Поскольку отрезки колеоптилей должны обладать одинаковой ростовой активностью (рост растяжением), нижняя часть колеоптиля, где рост растяжением уже практически окончен, также удаляется. Не снимая средние отрезки со станочка, из них при помощи тупой стеклянной иглы выталкивают и совершенно удаляют первичный лист. Получившиеся таким путем полые колеоптильные цилиндрики длиной 0,5 см нанизывают [c.18]
Новая стенка возникает сразу же после деления ядра из множества цитоплазматических пузырьков. Эти пузырьки сливаются с образованием тонкого диска (так называемой клеточной пластинки), который растет за счет все новых пузырьков, до тех пор пока не достигнет боковых стенок делящейся клетки. Цитоплазма двух дочерних клеток остается тесно связанной посредством большого числа тяжей цитоплазмы, которые пронизывают растущий диск, или пластинку. Тонкая клеточная пластинка быстро становится более основательной и вскоре приобретает форму первичной клеточной оболочки. Первичная оболочка была определена Уордропом [31 ] как структура, которая окружает протопласт в фазе роста клетки путем растяжения. Этому определению удовлетворяют клеточные стенки делящихся меристематических клеток, а также оболочки удлиняющихся клеток. Однако, хотя в обоих этих случаях мы имеем дело с первичными оболочками, [c.85]
Меристематические клетки обычно находятся в состоянии прекариотического равновесия. Клеточная оболочка поддеришвает это равновесие благодаря тому, что она ограничивает разрастание протопласта скорость роста клетки путем растяжения такова, что за время одного деления клетка увеличивается лишь вдвое. Однако, когда объем меристематической клетки растет с достаточно большой скоростью, скорость клеточного деления уменьшается. Для синтеза компонентов клеточной оболочки расходуется так много энергии, что достаточно каким-либо образом довести скорость роста до величины, хотя бы ненамного превышающей скорость, необходимую для поддержания постоянного размера клеток, и клетка уже оказывается более не в состоянии накапливать метаболиты, нужные для осуществления мито- [c.85]
Глава 31. Увеличение размеров клетки (рост в фазе растяжения). Дж. Лоета/7т 505 [c.623]
Следуя представлениям Бера (Вег, 1949), Хен считает, что в растущих клетках растений ауксины соединены с нуклеопротеидами и тиоурацил инактивирует этот комплекс. Урацил восстанавливает активность комплекса во время эмбрионального роста, но не в фазе роста растяжением, когда инактивация комплекса тиоурацилом носит иной характер (это способна сделать только ИУК). Автор предполагает, что для фазы роста растяжением характерна повышенная прочность связи тиоурацила с РНК. Возможно, что бор также играет какую-то важную роль в комплексировании ауксинов с нуклеопротеидами, а в его отсутствие этот комплекс инактивирован так же, как в присутствии тиоурацила, который вызывает такие же морфологические изменения, какие наблюдаются и при борной недостаточности (Heslop-Harrison, 1962). [c.119]
Количество ИУК в клетке в период медленного роста быстро увеличивается, достигая максимума 25 августа (рис. 6). Начало массового деления клеток связано с исключителыно резким уменьшением содержания ИУК- Дальнейшие изменения в содержании ИУК плоть до 8 сентября направлены в сторону некоторого повышения, хотя нужно отметить, что общий уровень ауксинов в клетке во время роста растяжением значительно 1НИ1же, чем в период, предшествующий началу массового деления клеток. [c.134]
Для роста клеток растяжением также небезразличен уровень кальция в клетке. Ингибитор кальмодулина Ы-(6-ами-ногексил)-5-хлор-1-нафтилсульфонамидгидрохлорид подавляет удлинение колеоптилей овса и кукурузы, а его неактивные аналоги не влияют на рост растяжением. Повышенная концентрация кальция тормозит ауксинзависимый рост, но усиливает способность ИУК индуцировать электрофизиологическую поляризацию тканей. Кальций — необходимый компонент в механизме поляризации клеток. [c.249]
Действие ауксина на рост растяжением сопровождается синтезом РНК, белков и секрецией полисахаридов и белков, необходимых для процесса клеточного растяжения. Рост клеточной стенки обеспечивается активной деятельностью АГ. Везикулы Гольджи поставляют полисахаридные компоненты, а также способствуют увеличению площади цитоплазматической мембраны. Усиливается синтез целлюлозы на наружной поверхности плазмалеммы. Выбор направления роста — удлинение клетки или ее изодиаметрический рост — определяется расположением микрофибрилл целлюлозы в клеточных стенках. Важно отметить, что к действию ауксина наиболее чувствителен эпидермис побегов. Рост внутренних паренхимных тканей сильнее активируется увеличением кислотности, вызванным, например, СО2 в повышенных концентрациях. Усиление роста под действием кислотности (pH 3 — 5) получило название эффекта кислого роста . Кислотность ускоряет рост лишь в течение [c.330]
На всех этапах роста клеток растяжением осуществляется самосборка работа генетического аппарата, обеспечивающего реализацию программы роста растяжением, взаимодействие ауксина с рецептором, сборка сложных ферментативных комплексов, в частности синтазы целлюлозы, состоящей из многих субъединиц, включение мембран пузырьков Гольджи в состав плазмалеммы растущей клетки, сборка молекул целлюлозы, гемицеллюлоз, пектинов и структурных белков (экстенсина) в клеточных стенках, процессы кристаллизации в микрофи-бриллах целлюлозы и т. д. [c.331]
Процессы самосборки осуществляются в течение всего жизненного цикла клетки, которая проходит этапы деления, роста растяжением, дифференцировки, старения и смерти. У делящихся растительных клеток нет центриолей, функцию которых выполняют скопления ретикулярных элементов. Деление клетки происходит благодаря образованию фрагмопласта (срединной пластинки). Для растений характерен рост клеток растяжением, механизм которого состоит в образовании большой центральной вакуоли и в размягчении и растяжении клеточной стенки. [c.333]
Рост и развитие стебля. Стержневая меристема апекса и прокамбий, образование которого индуцируется растущими примордиями листьев, формируют основные ткани стебля. Выходя из меристематической зоны, клетки начинают растягиваться что приводит к быстрому удлинению побега. Зона роста растя жением у побегов в отличие от корней достигает больших раз меров (нескольких сантиметров). Рост растяжением стебля ак тивируется гиббереллинами, стимулирующими переход боль шого количества клеток к этому типу роста, и ауксином который непосредственно индуцирует удлинение клеток. Гиббе реллины транспортируются из корня, но главным образом из листьев, и это позволяет регулировать скорость и продолжительность роста верхних междоузлий. [c.349]
В некоторых случаях продвижение в пространстве у растений достигается за счет верхушечного роста (гифы грибов, пыльцевые трубки, корневые волоски). С использованием изменений тургорного давления в клетках осуществляются движения устьиц, медленные настические движения листьев или быстрые движения листьев в ответ на сотрясение (сейсмонастии). Однако для подавляющего большинства растений, начиная с нитчатых водорослей, характерным способом перемещения в пространстве является рост растяжением. В основе движения растений лежат осмотические процессы в отличие от движений животных, происходящих с участием сократительных белков. [c.390]
Удлинение нити за счет деления клеток — процесс очень медленный. Появление у растительных клеток способности быстро удлиняться путем образования большой центральной вакуоли и растяжения клеточной стенки явилось приобретением, которое можно назвать ароморфозом. Действительно, для индивидуальной клетки ее удлинение и постенное расположение цитоплазмы оказались оптимальными для поглощения света хлоропластами. Нитчатая водоросль, удлиняющаяся за счет растяжения клеток, получила возможность гораздо быстрее двигаться к свету, поскольку длина клетки за сравнительно короткое время увеличивается в десятки и сотни раз. Такая форма движения по необходимости должна быть необратимой и поэтому одновременно является элементом морфогенеза. Удлинение клеток за счет роста рястяжением оказалось настолько удачной формой движения, что наряду с фотосинтезом стало основой развития растительного мира. Легко убедиться, что рост растяжением характерен только для растительных организмов ни у бактерий, ни у животных такого способа роста клеток не существует, так как рост растяжением возник у растений как способ движения многоклеточных автотрофных организмов (В. В. Полевой, Т. С. Саламатова, 1985). [c.411]
Главный способ движения органов растений связан с особым типом роста клеток — ростом растяжением. За счет зон роста растяжением в стебле и корне происходит более или менее интенсивное удлинение осевых органов в зависимости от условий окружающей среды (необратимый способ движения). Более обратимы reo-, фото-, хемо-и другие типы тропизмов (ростовых изгибов) и полностью обратимы настии, многие из которых осуществляются за счет изменения тургорного давления в клетках (тургорные изгибы). Возможно, все эти формы движений возникли из круговых нутаций, присущих всем растениям, особенно в молодом возрасте. Особый тип удлинения клеток — верхушечный рост — характерен для корневых волосков, пыльцевых трубок, гиф грибов. Регуляция ростовых и тургорных движений осуществляется с участием фитогормонов. [c.412]
Утолщение стеблей и корней происходит за счет делсиия клеток, роста растяжением и диффереицировки. У двудольных растений и у больщииства голосеменных радиальный рост стеблей и корней обеспечивается активностью латеральной меристемы. Обычно различают два типа латеральной меристемы сосудистый камбий и пробковый камбий, или феллоген. У тех видов-однодольных растений, которые в какой-то степени растут в толщину, клетки камбия также играют определенную, хотя и более ограниченную роль. [c.185]
Жизнь зеленого растения (1983) — [
c.72
,
c.82
,
c.84
,
c.92
,
c.259
,
c.260
,
c.273
,
c.276
]
Физиология растений (1989) — [
c.4
,
c.24
,
c.30
,
c.329
,
c.330
,
c.390
,
c.395
,
c.410
,
c.412
]
Источник