Рефлекс растяжения скелетной мышцы

Мышечные рефлексы (продолжение)
27 ноября 2012
Поделиться на:
![]()
Всем известно, что идеальной техникой бега обладают совсем маленькие дети, которые только что научились ходить. Но как им удается, не имея представления о том, что они делают, выполнять движения с наивысшим КПД. Кроме этого в грудном возрасте все люди крепко хватаются за пальцы — это рефлекс.
Интересно было наблюдать за сыном, когда он еще не умел ходить. Но когда я касался своими большими пальцами его ладошек, он так крепко их обхватывал, что я поднимал его из кроватки. А вот еще один интересный эксперимент, который мы с ним проделывали в то время. Я обхватывал его под мышками и удерживал над диваном так, что он едва касался пальцами ног покрывала. И он начинал выполнять беговые движения. Если при этом я проносил его вперед, то создавалось полное впечатление, что он бежит.
Мышечные рефлексы
В скелетных мышцах имеются два специализированных типа нервных рецепторов, которые могут воспринимать напряжение. Это мышечные веретена и сухожильный орган Гольджи (GTO’s). Мышечные веретена имеют сигарообразную форму и состоят из крошечных измененных мышечных волокон, называющихся интрафузалъными волокнами, и нервных окончаний, вместе покрытых соединительнотканной оболочкой. Они располагаются между и параллельно главным мышечным волокнам. Сухожильный орган Гольджи расположен главным образом в месте соединения мышц и их сухожилий или апоневроза.
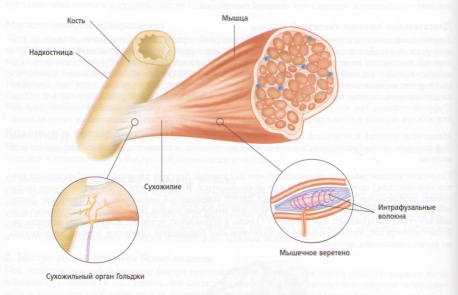
Рис. 18. Анатомия мышечного веретена и сухожильного органа Гольджи
Рефлекс растяжения мышцы
Рефлекс растяжения мышцы обеспечивает сохранение положения тела благодаря поддержанию мышечного тонуса. Он также предотвращает повреждение, давая возможность мышце отвечать на внезапное или неожиданное увеличение длины. Механизм рефлекса следующий:
1. Когда мышца удлиняется, мышечные веретена также растягиваются, заставляя каждое веретено посылать нервный импульс спинному мозгу.
2. При получении импульса спинной мозг немедленно посылает импульс обратно растянутым мышечным волокнам, заставляя их сокращаться, чтобы сопротивляться дальнейшему мышечному растяжению. Этот циркулярный процесс называется рефлекторная дуга.
3. Импульс одновременно посылается от спинного мозга до антагониста мышечного сокращения (т. е. мышце, выступающей против сокращения), вызывая расслабление антагониста, чтобы он не мог сопротивляться сокращению растянутого мышечного волокна. Этот процесс называется реципрокное торможение.
4. Параллельно этому спинальному рефлексу нервные импульсы также посылаются от спинного к головному мозгу с целью передачи информации относительно длины мышцы и скорости ее сокращения. Головной мозг посылает обратно нервные импульсы к мышце для поддержки соответствующего мышечного тонуса, чтобы обеспечить требуемое положение тела и движения.
5. Тем временем чувствительность к растяжению мелких интрафузальных мышечных волокон в пределах мышечного веретена сглаживается и регулируется гамма-эфферентными нервными волокнами, отходящими от двигательных нейронов в спинном мозге. Таким образом, гамма-мотонейрон-рефлекторная дуга обеспечивает равномерность сокращения мышечной ткани, которое иначе было бы хаотичным, если бы мышечный тонус основывался на одном рефлексе растяжения.
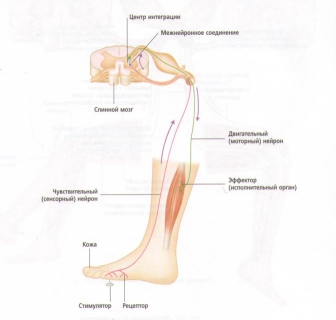
Рис. 19. Основная рефлекторная дуга.
На практике классическим клиническим примером рефлекса растяжения мышцы является коленный рефлекс, или коленный маятникообразный рефлекс, когда по надколенному сухожилию слегка ударяют небольшим резиновым молотком. Рефлекс заключается в следующем:
1. Внезапный удар по надколенному сухожилию приводит к растяжению четырехглавой мышцы.
2. Быстрое растяжение регистрируется мышечными веретенами четырехглавой мышцы, вызывая ее сокращение. Это вызывает небольшой толчок, т. е. внезапное распрямление колена и снятие напряжения мышечных веретен.
3. Одновременно подавляются нервные импульсы к мышцам задней группы бедра, которые являются антагонистами четырехглавой мышцы, это приводит к расслаблению мышц задней группы бедра.
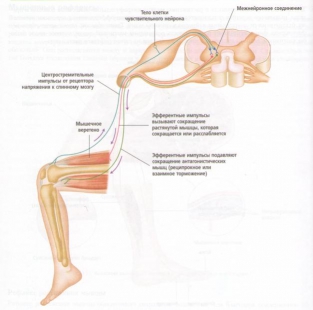
Рис. 20. Рефлекторная дуга напряжения.
На практике другим ярким примером рефлекторной дуги растяжения является следующее: когда человек засыпает в сидячем положении, голова наклоняется вперед, затем отдергивается, потому что напряженные мышечные веретена задней части шеи активизировали рефлекторную дугу.
Рефлекс растяжения мышцы работает постоянно для поддержания тонуса постуральных мышц (т. е. мышц, обеспечивающих правильную осанку). Таким образом рефлекс позволяет человеку оставаться в вертикальном положении без сознательных усилий и без падения вперед. Последовательность событий, предотвращающих падение вперед, происходит за доли секунды следующим образом:
1. В вертикальном положении человек естественно начинает покачиваться вперед.
2. Икроножные мышцы растягиваются, активизируя рефлекс напряжения.
3. Икроножные мышцы последовательно сокращаются, удерживая человека в вертикальном положении.
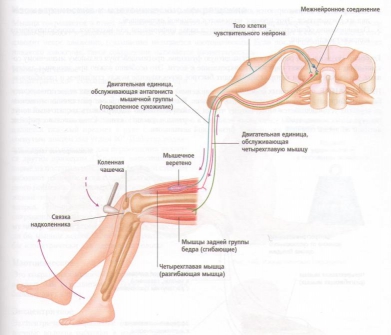
Рис. 21. Коленный маятникообразный рефлекс.
Глубокий сухожильный рефлекс (аутогенное торможение)
В противоположность рефлексу растяжения, который вызывает реакцию мышечного веретена, глубокий сухожильный рефлекс вызывает реакцию сухожильных органов Гольджи, приводящую к мышечному сокращению. Глубокий сухожильный рефлекс вызывает противоположный эффект рефлексу растяжения. Он работает следующим образом:
1. При сокращении мышцы происходит натягивание сухожилий, расположенных с обоих концов мышцы.
2. Напряженность в сухожилии заставляет сухожильный орган Гольджи передавать импульсы спинному мозгу (большое количество импульсов передается мозжечку).
3. Как только импульсы достигают спинного мозга, они подавляют двигательные нервы, обеспечивающие сокращение мышцы, вызывая расслабление.
4. Одновременно активизируются двигательные нервы, обслуживающие мышцы-антагонисты, заставляя их сокращаться. Этот процесс называется взаимной активацией.
5. Одновременно обрабатывается и отправляется назад информация для мозжечка, корректирующего напряженность мышц.
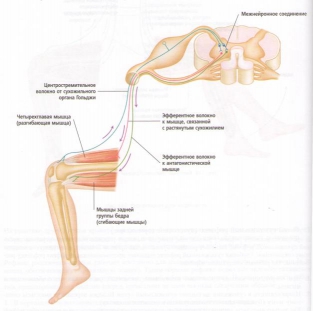
Рис. 22. Глубокий сухожильный рефлекс.
Глубокий сухожильный рефлекс имеет защитную функцию, противодействуя сильному мышечному сокращению, которое может нарушить прикрепление к кости. Это особенно важно при активных движениях, таких, как бег, при которых происходит быстрое переключение между сгибанием и разгибанием.
ПРИМЕЧАНИЕ: при обычных повседневных движениях напряженности в мышцах недостаточно, чтобы активизировать глубокий сухожильный рефлекс. В отличие от этого порог растяжения мышечного веретена устанавливается ниже, потому что он должен постоянно поддерживать достаточный тонус постуральных мышц, чтобы удерживать человека в вертикальном положении. Следовательно, рефлекс активен во время обычной ежедневной физической активности
Нам осталось рассмотреть еще один не маловажный, а даже наоборот — основополагающий вопрос. Это конечно, питание. И связанные с ним мероприятия восстанивления. Не знаю будет ли под силу в одной статье охватить столь жизненный и повседневный вопрос.
Источник
Мышечные волокна
Скелетные мышцы состоят из пучков мышечных волокон. Мышечные волокна могут сокращаться, расслабляться или удлиняться.
Мышечные волокна сокращаются под действием электрического импульса, поступающего от нерва. Одно мышечное волокно всегда сокращается полностью, создавая фиксированную силу. Сила, развиваемая всей мышцей, определяется количеством волокон, вовлеченных в это действие нервной системой.
Соединительные ткани
Соединительные ткани в опорно-двигательном аппарате — это связки, сухожилия и фасции. Связки обоими концами крепятся к костям сустава, сухожилия крепят мышцы к костям, а фасции обволакивают группы мышц и пучки мышечных волокон.
Связки и сухожилия содержат большое количество белка коллагена, который придает им жесткость. Связки и сухожилия практически не растягиваются — избыточная нагрузка на эти ткани может привести к травме. Фасции относятся к эластичным соединительным тканям (содержат белок эластин), и их растягивание повышает гибкость. Большинство упражнений на гибкость направлено на растяжение фасций.
Механизм растягивания
При растягивании сначала вытягиваются мышечные волокна, а затем в направлении вытягивающего усилия выравниваются коллагеновые волокна соединительной ткани. Это позволяет упорядочить волокна в направлении растяжения, благодаря чему в упражнениях на растяжение восстанавливается здоровая структура ткани.
При растяжении мышц часть мышечных волокон растягивается, а часть остается в исходном состоянии. Длина мышцы определяется количеством растянутых волокон (аналогично тому, как длина и сила сжатой мышцы определяется количеством сжатых волокон). Чем больше растянутых волокон, тем длиннее растянутая мышца.
Рефлекс растяжения
В толще мышц находятся нервно-мышечные веретена — нервные окончания, реагирующие на удлинение мышц. Веретена располагаются параллельно мышечным волокнам и растягиваются вместе с ними. При этом они регистрируют как изменение длины мышцы, так и скорость этого изменения.
При сильном или резком удлинении мышцы сигнал от веретен вызывает защитный рефлекс растяжения — мышца самопроизвольно сокращается в попытке помешать удлинению.
Рефлекс растяжения имеет статическую и динамическую составляющие. Статическая составляющая сохраняется все время, пока мышца растянута. Динамическая составляющая — это ответ на скорость изменения длины мышцы, он может быть очень сильным при резком удлинении мышцы и снижается, когда скорость удлинения мышцы уменьшается.
Одна из целей удержания позы растяжения в течение некоторого времени — дать мышечным веретенам адаптироваться к новой длине мышцы с уменьшением интенсивности их сигналов в новом положении.
Реакция удлинения
При сокращении мышца создает натяжение на сухожилии, к которому она крепится. На это реагирует сухожильный орган Гольджи — тип нервного окончания, который находится в местах соединения волокон мышц и сухожилий. Сухожильный орган реагирует на величину возникающей силы натяжения (статический эффект) и на скорость изменения этой силы (динамический эффект: резкое натяжение вызывает сильный сигнал).
Когда сигнал от сухожильного органа превосходит определенный порог, возникает защитная реакция удлинения — резкое самопроизвольное расслабление всей мышцы.
Вторая причина для удержания позы растяжения в течение некоторого времени — позволить произойти реакции удлинения, которая помогает мышце расслабиться (преодолевая рефлекс сжатия).
Взаимное торможение
По отношению к выполняемому движению мышцы можно отнести к одному из четырех типов:
- Агонисты — вызывают движение.
- Антагонисты — вызывают противоположное движение; отвечают за возвращение в начальное положение тела.
- Синергисты — корректируют действие мышцы-агониста для обеспечения нужного направления результирующей силы.
- Стабилизаторы — удерживают остальную часть тела при выполнении движения.
Агонисты и антагонисты обычно находятся с противоположных сторон сустава (например, бицепс и трицепс), а синергисты — на той же стороне, что и агонисты, рядом с ними. При работе крупных мышц-агонистов часто вовлекаются в качестве синергистов находящиеся рядом меньшие мышцы.
Сокращение мышцы-агониста может привести к рефлекторному расслаблению ее антагониста. Это явление называется взаимным торможением.
Взаимное торможение происходит не при всех движениях. Иногда возникает явление совместного сжатия. Например, при приседаниях сжимаются как мышцы брюшного пресса, так и их антагонисты — разгибатели спины.
Упражнения на растяжения легче выполнять с расслабленными, а не с сокращающимися мышцами. Для этого можно использовать эффект взаимного торможения (когда он происходит) — заставляя мышцы-антагонисты расслабляться за счет сжатия агонистов. Также желательно расслабить синергистов мышцы, которую вы растягиваете. Например, при растягивании икроножной мышцы, следует нагнуть стопу, сжав мышцы передней поверхности голени. С другой стороны, икроножная мышца — синергист подколенного сухожилия, поэтому желательно расслабить и его. Для этого нужно выпрямить ногу, сжав тем самым антагониста подколенного сухожилия — четырёхглавую мышцу бедра.
Источник
Реакция первичного окончания мышечного веретена. Рефлекс на растяжение мышцыКогда длина мышечного веретена изменяется внезапно, первичное окончание (но не вторичное) стимулируется особенно мощно. Это избыточное возбуждение первичного окончания называют динамическим ответному что означает чрезвычайно активную реакцию первичного окончания на высокую скорость изменения длины веретена. Даже когда длина веретена увеличивается лишь на долю микрометра и это увеличение осуществляется в течение доли секунды, первичный рецептор передает громадное количество дополнительных импульсов по большим сенсорным нервным волокнам диаметром 17 мкм, но только до тех пор, пока длина действительно увеличивается. Как только увеличение длины прекращается, этот дополнительный прирост импульсного разряда возвращается к гораздо более низкому уровню, чем статический разряд, все еще присутствующий в ответе. Наоборот, при укорочении веретена происходит прямо противоположное изменение сигнала. Таким образом, первичное окончание посылает чрезвычайно сильные, положительные или отрицательные, сигналы к спинному мозгу, информируя его о любом изменении длины мышечного веретена. Регуляция интенсивности статических и динамических ответов гамма-мотонейронами. Гамма-двигательные нервы к мышечному веретену можно разделить на два типа: гамма-динамические (гамма-d) и гамма-статические (гамма-s). Первые из них возбуждают главным образом интрафузальные волокна с ядерной сумкой, а вторые возбуждают в основном интрафузальные волокна с ядерной цепочкой. Когда гамма-d волокна возбуждают волокна с ядерной сумкой, динамический ответ мышечного веретена становится чрезвычайно усиленным, тогда как статический ответ почти не меняется.
Наоборот, стимуляция гамма-s волокон, возбуждающих мышечные волокна с ядерной цепочкой, усиливает статический ответ, оказывая лишь незначительное влияние на динамический ответ. Непрерывный разряд мышечных веретен при нормальных условиях. В норме, особенно на фоне некоторой степени возбуждения гамма-эфферентных волокон, в сенсорных нервных волокнах мышечных веретен постоянно возникают импульсы. Растяжение мышечных веретен увеличивает частоту импульсов, тогда как укорочение веретен снижает ее. Таким образом, веретена могут посылать к спинному мозгу положительные сигналы, т.е. увеличенное количество импульсов, свидетельствующее о растяжении мышцы, или отрицательные сигналы, т.е. количество импульсов ниже нормы, что указывает на отсутствие растяжения мышцы. Рефлекс на растяжение мышцыСамым простым проявлением функции мышечных веретен является рефлекс на растяжение мышцы. Всякий раз, когда мышца внезапно растягивается, возбуждение веретен вызывает рефлекторное сокращение крупных мышечных волокон растянутой мышцы и тесно связанных с ней мышц-синергистов. Нервный контур рефлекса на растяжение. На рисунке изображен основной контур рефлекса на растяжение мышечных веретен. Видно, что проприоцептивное нервное волокно типа 1а, исходящее от мышечного веретена, входит в задний корешок спинного мозга. Затем ветвь этого волокна идет непосредственно к переднему рогу серого вещества спинного мозга и синаптически связывается с передними мотонейронами, которые посылают двигательные нервные волокна к той же мышце, от которой исходят волокна мышечных веретен. Таким образом, существует моносинаптический путь, позволяющий рефлекторному сигналу вернуться с кратчайшей задержкой назад к мышце после возбуждения веретена. Большинство волокон типа II от мышечных веретен заканчиваются на множестве вставочных нейронов серого вещества, и их аксоны проводят сигналы к передним мотонейронам с задержкой или выполняют другие функции. — Также рекомендуем «Динамические и статические рефлексы на растяжение. Демпфирующий механизм сокращения» Оглавление темы «Физиология спинальной рефлекторной деятельности»: |
Источник
В активной части опорно двигательного аппарата имеются проприорецепторы. Существует два типа проприорецепторов – нервно-мышечные веретёна, которые реагируют на изменение длины мышцы, а также сухожильные органы Гольджи, реагирующие на изменение напряжения, то есть на силу сокращения мышечных волокон (Рисунок 1).

Рисунок 1. Проприорецепторы: нервно-мышечное веретено и сухожильный орган Гольджи
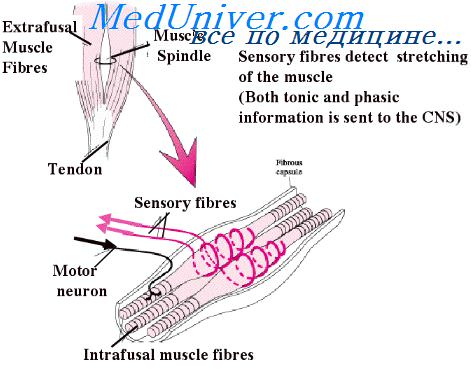
Нервно-мышечные веретёна. Нервно-мышечные веретена расположены в толще скелетных мышц между мышечными волокнами. Они представляют собой веретеновидные структуры, покрытые соединительнотканной капсулой. Длина веретена составляет 4 – 7 (до 10) мм, диаметр – 100 мкм. Под капсулой внутри веретена находятся видоизменённые мышечные волокна, которые называются интрафузальными. Эти волокна имеют ряд отличий от обычных, экстрафузальных, мышечных волокон. Во-первых, интрафузальное волокно имеет меньший диаметр (15 – 30 мкм) по сравнению с экстрафузальным (50 – 100 мкм). Во-вторых, в центральной («экваториальной») части интрафузального волокна нет сократительных элементов, поэтому к сокращению способны только периферические части. Поскольку концы интрафузального волокна фиксированы к соединительнотканной капсуле, покрывающей веретено, при укорочении периферических отделов волокна его центральная часть растягивается. Интрафузальные волокна расположены параллельно эктсрафузальным. По строению интрафузальные волокна делятся на два типа – с ядерной сумкой и с ядерной цепочкой (Рисунок 2). В свою очередь, волокна с ядерной сумкой по функции по особенностям иннервации подразделяются на статические и динамические. Для волокон с ядерной сумкой характерно наличие расширенной центральной части, в которой ядра собраны в скопления. Волокна с ядерной цепочкой примерно вдвое тоньше волокон с ядерной сумкой, ядра в их центральной части расположены в ряд. В каждом веретене содержится 1 – 3 волокна с ядерной сумкой и 5 – 7 волокон с ядерной цепочкой.
Рисунок 2. Нервно-мышечное веретено
Интрафузальные волокна получают как афферентную, так и эфферентную иннервацию. Выделяют два типа афферентных окончаний – первичные и вторичные. Первичные окончания называют аннулоспиральными, они представляют собой спираль, окружающую центральную часть всех интрафузальных волокон. От первичных окончаний отходят волокна типа Ia, которые соответствуют типу Aα по Гассеру-Эрлангеру. Это самые толстые волокна (~17 мкм) с самой высокой скоростью проведения возбуждения (70 – 12 м/с). Каждое нервно-мышечное веретено иннервируется только одним волокном Ia. Другими словами одно афферентное нервное волокно Ia получает импульсацию со всех первичных окончаний в иннервируемом им веретене. Вторичные окончания имеют форму спирали, но могут также образовывать разветвлённую структуру. Они оплетают центральную часть интрафузальных волокон с ядерной цепочкой, а также статических волокон с ядерной сумкой, но их не бывает в динамических волокнах с ядерной сумкой. От вторичных окончаний отходят волокна типа II, соответствующие типу Аβ по Гассеру-Эрлангеру; диаметр этих волокон составляет ~8 мкм, они проводят возбуждение со скоростью 40 – 70 м/с (Рисунок 3).

Рисунок 3. Схема нервно-мышечного веретена
Как первичные, так и вторичные нервные окончания чувствительны к изменению длины интрафузальных волокон, однако характер ответной реакции различается. При медленном растяжении центральной части интрафузальных волокон импульсация появляется как в первичных, так и во вторичных рецепторах. Частота импульсов пропорциональная степени растяжения волокна. Такой ответ афферентных окончаний называют статическим. Однако при быстром внезапном растяжении волокна в первичных (но не вторичных) окончаниях отмечается мощный всплеск импульсации – динамический ответ. Следовательно, волокна Ia передают информацию как об абсолютной длине интрафузальных волокон (то есть о степени растяжения), так о скорости её изменения, тогда как волокна II – только об абсолютной длине. Следует понимать, что растяжение центральной части интрафузальных волокон может быть связано как с удлинением всей мышцы (интрафузальные волокна расположены параллельно экстарфузальным), так и при сокращении периферических отделов волокна (Рисунок 3).

Рисунок 4. Статический и динамический ответы рецепторов нервно-мышечных веретён
Эфферентная иннервация интрафузальных волокон представлена нервными волокнами типа Aγ, которые являются аксонами γ-мотонейронов спинного мозга. Диаметр этих волокон составляет ~5 мкм, скорость проведения возбуждения в них – 15 – 40 м/с. Окончания Aγ волокон подходят к периферическим отделам интрафузальных волокон, при возбуждении γ-мотонейронов концевые участки интрафузальных волокон сокращаются, в результате чего их центральные отделы растягиваются. Выделяют γ-динамические и γстатические волокна, γ-динамические волокна иннервируют динамичекие волокна с ядерной сумкой, их активация повышает динамическую чувствительность окончаний Iа типа.
γ-статические волокна иннервируют волокна с ядерной цепочкой и статические волокна с ядерной сумкой, их возбуждение повышает уровень активности как первичных, так и вторичных окончаний (см. ниже).
Миотатический рефлекс (рефлекс растяжения)
В ответ на увеличение длины мышцы рецепторы нервно-мышечных веретён возбуждаются, и импульсация по волокнам Ia (и при быстром, и при медленном растяжении) или II (при медленном растяжении) поступает в спинной мозг. Войдя в серое вещество спинного мозга, волокно афферентного нейрона разветвляется и образует возбуждающие синапсы практически со всеми α-моторными нейронами, иннервирующими гомонимную (то есть эту же) мышцу. Возбуждение α-моторных нейронов вызывает сокращение экстрафузальных волокон, и мышца укорачивается. Следует отметить, что коллатерали афферентных нейронов переключаются также на вставочные тормозные нейроны, которые угнетают α-мотонейроны мышц антагонистов. Таким образом, растяжение мышцы вызывает её рефлекторное укорочение, при этом мышцы антагонисты расслабляются. Описанный рефлекс с рецепторов нервно-мышечных веретён называется миотатическим, или рефлексом на растяжение (Рисунок 4). Миотатический рефлекс является моносинаптическим, поэтому имеет очень небольшой латентный период (~ 0,9 мс).

Рисунок 5. Миотатический рефлекс с нервно-мышечных веретён
Различают фазический (динамический) и тонический миотатический рефлекс. Фазический рефлекс запускается с первичных нервных окончаний при быстром растяжении мышцы, например, при ударе по сухожилию неврологическим молоточком или при появлении внезапного препятствия совершаемому движению. Тонический рефлекс запускается с вторичных нервных окончаний при медленном (или постоянном) растяжении мышечных волокон; он имеет значение для поддержания вертикальной позы. Когда тело отклоняется от правильного положения, мышцы, участвующие в поддержании позы, растягиваются. В результате возбуждаются как первичные, так и вторичные окончания нервно-мышечных веретён, запускается миотатический рефлекс, и мышцы сокращаются; за счёт их сокращения тело возвращается в правильное положение. Когда у длительно стоящего человека (например, солдата в стойке «смирно») из-за утомления сгибаются ноги в коленных суставах, проприорецепторы четырёхглавых мышц реагируют на растяжение, и ноги снова выпрямляются. Постоянное стимулирующее влияние на α-мотонейроны со стороны афферентных волокон Ia и II типов вносит вклад в поддержание мышечного тонуса.
Функции γ-моторных нейронов. Понятие о γ-петле.
Как уже упоминалось, интрафузальные волокна получают эфферентную иннервацию со стороны γ-моторных нейронов. При возбуждении γ-мотонейронов происходит сокращение периферических (концевых) отделов интрафузальных волокон, в результате их центральная часть растягивается. При этом первичные и вторичные нервные окончания активируются, импульсация от них поступает в спинной мозг, и возбуждаются α-моторные нейроны. Другими словами, запускается миотатический рефлекс. Такой механизм (активация рефлекса растяжения при возбуждении γ-мотонейронов) называется γ-петлёй. Наличие γ-петли обеспечивает сохранение чувствительности рецепторов нервномышечных веретён при разной длине мышцы. Во время сокращения мышцы её длина уменьшается, соответственно должны укорачиваться и интрафузальные волокна. В результате импульсация с рецепторов нервно-мышечных веретён должна была бы прекратиться, и рецепторы потеряли бы способность реагировать на изменение длины мышцы. Напротив, при расслаблении мышцы её длина увеличивается, и интрафузальные волокна могут быть чрезмерно растянуты. В таких условиях рецепторы будут генерировать импульсы с максимальной частотой и, следовательно, также потеряют способность реагировать на изменение длины. Однако в реальных условиях имеет место α-γ-коактивация, то есть α- и γмотонейроны возбуждаются одновременно. В результате сокращаются как экстра-, так и интрафузальные волокна. Сокращение экстрафузальных волокон приводит к укорочению мышцы, а сокращение периферических отделов интрафузальных волокон – к растяжению их центральной части. Следовательно, длина центральной части не меняется, поскольку её укорочение вследствие сокращения экстрафузальных волокон компенсируется растяжением из-за сокращения периферических отделов интрафузальных волокон. И наоборот – расслабление всей мышцы сопровождается расслаблением и интрафузальных волокон, при этом растяжение центральной части уменьшается. Таким образом, за счёт α-γ-коактивации рецепторы нервно-мышечных веретён сохраняют чувствительность (и, следовательно, передают информацию о длине мышцы в ЦНС) как при сокращении мышцы, так и при её расслаблении (Рисунок 6). На γ-мотонейронах заканчиваются нисходящие пути из различных отделов головного мозга. Меняя активность γ-мотонейронов, мозговые центры влияют на чувствительность рецепторов нервно-мышечных веретён и, следовательно, на порог рефлекса растяжения. Сухожильные органы Гольджи. Обратный миотатический рефлекс Кроме нервно-мышечных веретён в активной части опорно-двигательного аппарата имеется ещё один тип проприрецепторов – сухожильные органы Гольджи. Сухожильные органы представляют собой сеть коллагеновых нитей, окружённую тонкой соединительнотканной капсулой. Диаметр сухожильного органа Гольджи составляет 100 мкм, длина – 1 мм. Коллагеновые нити внутри органа оплетённых нервными окончаниями, от которых отходят нервные волокна типа Ib. эти нервные волокна, как и Ia, соответствуют типу Aα по Гассеру-Эрлангеру. Сухожильные органы Гольджи расположены в месте прикрепления мышечных волокон к сухожильным, при этом каждый сухожильный орган связан с 3 – 25 (в среднем – с 10 – 15) мышечными волокнами (Рисунок 7). Таким образом, в отличии от нервно-мышечных веретён, которые ориентированы параллельно мышечным
волокнам, сухожильные органы по отношению к мышечным волокнам расположены последовательно.

Рисунок 6. Функция γ-петли
Следовательно, сухожильные органы реагируют не на изменение длины мышцы, а на развиваемое ей усилие, другими словами, на степень её напряжения. Афферентные нервные волокна от сухожильных органов Гольджи следуют в спинной мозг, где, в отличие от чувствительных волокон с нервно-мышечных веретён, переключаются на вставочные тормозные нейроны. Вставочные нейроны, в свою очередь, образуют синаптические контакты с α-мотонейронами гомонимной мышцы. Таким образом, активация сухожильных органов приводит к расслаблению мышцы, к которой крепится данное сухожилие. Описанный рефлекс называется обратным рефлексом растяжения (обратным миотатическим рефлексом) (Рисунок 8). Этот рефлекс, по-видимому, имеет две функции. Во-первых, он предупреждает чрезмерное напряжение скелетных мышц, которое может привести к тяжёлым травмам (разрыву мышцы, отрыву сухожилия от кости или от мышцы). Во-вторых, обратный миотатический рефлекс обеспечивает равномерность силы сокращения разных мышечных волокон в пределах одной мышцы. Те волокна, которые напряжены слишком сильно, расслабляются за счёт усиления тормозных влияний со стороны спинного мозга. Напряжение волокон, которые сокращаются недостаточно сильно, напротив, усиливается за счёт уменьшения тормозного воздействия.

Рисунок 7. Сухожильный орган Гольджи
Проприорецепторы посылают в головной мозг информацию о состоянии скелетно-мышечной системы Импульсация с проприорецепторов поступает не только в спинной мозг, но и по коллатералям афферентных волокон в различные отделы головного мозга. В частности, импульсы с нервно-мышечных веретён и сухожильных органов следуют по спинномозжечковым трактам в мозжечок, а также по системе задних столбов (тонкий и клиновидный пучок) в продолговатый мозг, затем – в таламус, а оттуда – в кору больших полушарий. Таким образом, мозжечок и кора большого мозга постоянно получают информацию о состоянии скелетно-мышечной системы, а именно о длине мышц и о степени их напряжения. Эта информация является необходимой как для регуляции произвольных движений и позы, так и для осознания положения частей тела в пространстве и друг относительно друга.

Рисунок 8. Обратный миотатический рефлекс с сухожильных органов Гольджи
Источник