Деление клеток при растяжении у человека

Стволовые клетки эпидермиса с помощью внутриклеточных скелетных белков чувствуют механическое напряжение и начинают особенно активно делиться.
Если наша кожа чувствует механическое напряжение, она увеличивается. То есть может показаться, что она растягивается, но на самом деле в ней просто становится больше клеток. Собственно, о том, что кожу можно заставить расти, растянув её, было известно давно, и это её свойство использовали и используют в медицине. Но как именно механическое напряжение переводится на язык межклеточных сигналов, мы до сих пор представляем себе плохо.
Кожа человека: над дермой (розовый толстый слой, занимающий большую часть фото) лежит базальный слой эпидермиса с углублениями в дерму. Стволовые клетки базального слоя обновляют верхние слои эпидермиса, которые постепенно отмирают. (Фото: Kilbad / Wikipedia)
‹
›
Очевидно, что тут главную роль играют стволовые клетки эпидермиса, благодаря которым кожа постоянно обновляется. Если кожа чувствует натяжение, стволовые клетки должны активнее делиться, причём часть дочерних клеток должна стать специализированной, то есть превратиться в рабочие клетки кожи, а часть должна так и остаться в стволовом состоянии.
Исследователи из Брюссельского свободного университета вживили мышам под кожу приспособление, которое используется в хирургии и которое медленно растягивает кожу, понуждая её расти; стволовые клетки кожи у мышей модифицировали генетически так, чтобы они синтезировали светящийся белок, по которому их и их потомков можно было бы легко узнать. Так удалось показать, что стволовые клетки в ответ на натяжение действительно начинают активнее делиться, и благодаря приросту стволовых клеток в коже появляется больше клеток обычных, из которых кожа преимущественно и сделана.
На уровне генов и молекул происходило следующее. Во-первых, активировались гены, от которых зависят межклеточные контакты – очевидно, что белки межклеточных контактов в первую очередь ощущают физическое напряжение при растяжении. Во-вторых, активировались гены, кодирующие белки цитоскелета, то есть внутренней белковой сети, которая служит клетке опорой, структурирует её и помогает двигаться. В цитоскелет входят разные белки, и при напряжении особенно много становилось тех, которые способны сокращаться и стягивать разные участки клетки. В-третьих, активировались гены сигнальной молекулярной цепочки, которая стимулирует клеточное деление. Всё это сопровождалось перестройками в хроматине, то есть в белково-хромосомном комплексе – нужные гены распаковывались и становились доступны для молекулярных машин, считывающих генетическую информацию.
В статье в Nature говорится, что не все стволовые клетки, которые попали под растяжение, реагировали подобным образом – то есть активировали гены для поддержания клеточных контактов клеточного деления и пр. Возможно, тут всё зависит от того, какая сила действует на клетку: есть клетки, которых растягивает сильнее, есть те, которых растягивает слабее.
С другой стороны, сами клетки могут отличаться чувствительностью к механическому напряжению, пусть даже объективно оно одинаково для всех – кто-то его будет ощущать сильнее, кто-то слабее. Чтобы ответить на этот вопрос, как пишет портал Nature, нужно измерить силы, действующие на отдельную живую клетку, что до сих пор остаётся очень сложной задачей.
Когда мышам вносили мутации в гены, кодирующие сократительные элементы цитоскелета, стволовые клетки в прямом смысле переставали чувствовать механическое напряжение, и кожа в ответ на растяжение не росла. Дальнейшие эксперименты показали, что сократительный цитоскелет, когда его растягивают, помогает проникнуть в клеточное ядро особым белкам, которые в ядре включают гены, необходимые для ответа на растяжение.
Иными словами, белковые сократительные нити в стволовой клетке чувствуют натяжение и переводят это ощущение (если можно говорить про ощущение у белков) на язык молекулярных сигналов. Реакция на натяжение включается не у всех стволовых клеток, однако, скорее всего, другие стволовые клетки чувствуют активность своих соседей и как-то на неё реагируют.
Полученные результаты пригодятся не только в медицине, где часто бывает нужно усилить рост кожи, чтобы закрыть рану или ожог. Механические силы вообще играют большую роль в жизни самых разных клеток, начиная от иммунных (в прошлом году мы писали, как механические силы регулируют активность иммунитета в лёгких) до опухолевых, которые давят на здоровые клетки вокруг себя и тем самым понуждают их к суициду. Расшифровав молекулярные механизмы, включающиеся в ответ на растяжение в стволовых клетках кожи, мы, возможно, лучше поймём, как управлять жизнью других наших клеток.
Источник
В тканях, подвергающихся длительному растягиванию, происходят значительные биологические и морфологические изменения. Несмотря на то, что площадь поверхности во время растягивания всегда увеличивается, покрывающий эпидермис не утончается, а, как было обнаружено, немного утолщается. Исследования показали, что митотическая активность растягиваемой кожи увеличивается, и увеличенный темп митозов при этом способствует сохранению и даже увеличению высоты эпидермиса при сохранении нормальной многослойной структуры. Изменения в эпидермисе представляются временными, и микроскопически кожа возвращается к норме в течение года или двух лет после растягивания. Клинически наиболее частыми видимыми изменениями кожи являются высыхание и гиперпигментация.
Однако дерма не отвечает на хроническое растягивание так же благоприятно, как эпидермис. Наблюдается значительное истончение дермы (на 30-50%), особенно при ускоренном растягивании. Во многих клеточных популяциях дермы отмечается увеличение метаболической активности. Активность фибробластов обычно увеличивается, и нарастает синтез коллагена. Более того, изменяется физическое расположение коллагеновых волокон, а эластические волокна фрагментируются. При растягивании может произойти увеличение пигментации, связанное с временным стимулированием выработки меланина. Волосяные луковицы становятся менее частыми из-за увеличения площади поверхности, но истинное число фолликулов остается прежним.
К тому же увеличивается толщина базального слоя и обнаруживается большее число миофибробластов, чем в нерастянутой коже. Эти изменения в дерме и придатках кожи могут вызывать клинические изменения, выражающиеся в жесткости, образовании полос (стрий), эритродермии, облысении, уменьшении чувствительности.
Процесс растягивания также затрагивает связанные структуры подкожных тканей. Жировая ткань очень плохо переносит растягивание и значительно истончается, теряя до 50% первоначальной толщины. Жировые клетки уплощаются, теряют жировые включения и могут заместиться фиброзной тканью. Однако эта утрата жира может быть до некоторой степени восполнена после растягивания. Сосудистые изменения при этом могут быть разительными.
Хроническое растягивание ткани является сильным стимулом пролиферации сосудов. Это одно из больших достоинств растягивания, так как этот процесс обычно приводит к улучшению кровоснабжения растягиваемого лоскута, что может быть особенно полезно в случаях, когда сосудистая сеть нарушена. Тогда растягивание может дать не только увеличение площади поверхности, но также улучшение кровоснабжения и большую устойчивость растянутой ткани к инфекции. При растягивании имеет место пролиферация капилляров и, разумеется, венул и артериол, а также удлинение сосудистых структур.
Клиническая практика показала, что растянутые лоскуты имеют, несомненно, более высокую частоту выживания, подобно отсроченным лоскутам. Вокруг баллона-расширителя (экспандера) неизменно образуется плотная фиброзная капсула, которая не только содержит много фибробластов, но и хорошо кровоснабжается (рис. 1). Капсула выстлана макрофагами, окруженными фибробластами, которые активно вырабатывают коллаген. Капсула может частично способствовать улучшению кровоснабжения растянутых лоскутов, но она также участвует в контрактуре и сжатии лоскута после удаления расширителя и использования лоскута по назначению. Эта последующая контрактура может вызвать утолщение лоскута и сделать его неподходящим для определенных реконструкций, требующих тонких, пластичных лоскутов (например, лобные лоскуты для реконструкции носа) (рис. 2). По необходимости капсулу можно иссечь предположительно без нарушения кровоснабжения лоскута в целом, хотя такое иссечение нужно выполнять осторожно.

Рис. 1. Лоскуты, сформированные после длительного растягивания тканей лба. Обратите внимание на толщину лоскутов и капсулу на их внутренней поверхности.

Рис. 2. Утолщившийся и сокращенный парамедиальный лобный лоскут, который растягивался перед формированием и перемещением на нос. Иссечение капсулы может помочь избежать этого.
Мышечная ткань не переносит растягивания так же хорошо, как сосудистая ткань. Наблюдается истончение, некроз, атрофия и клинически значимое ослабление мышцы. На голове и шее это может стать наибольшей проблемой для лобной мышцы и других мимических мышц. Нервная ткань лучше воспринимает растягивание, так что постепенное удлинение нервов может происходить без их некроза.
Экспериментальные исследования показали, что периферические нервы можно удлинять при длительном растягивании на 32%, но с некоторым ослаблением функции. Лицевой нерв в условиях быстрого растягивания изучался Martini и соавт., которые обнаружили, что лицевой нерв кота можно значительно удлинить путем интраоперационного растягивания. Однако 40% растянутых нервов не смогли регенерировать, вероятно, из-за механического разрыва аксонов или нарушения кровоснабжения при быстром интраоперационном растягивании. Большой проблемой растягивания тканей головы и шеи является то, что экспандер может ослабить мышцы лица и ветви лицевого нерва. Поэтому, возможно, для уменьшения риска разумно помещать растягивающие устройства на лице и шее поверхностнее SMAS и подкожной мышцы.
Опубликовал Константин Моканов
Источник
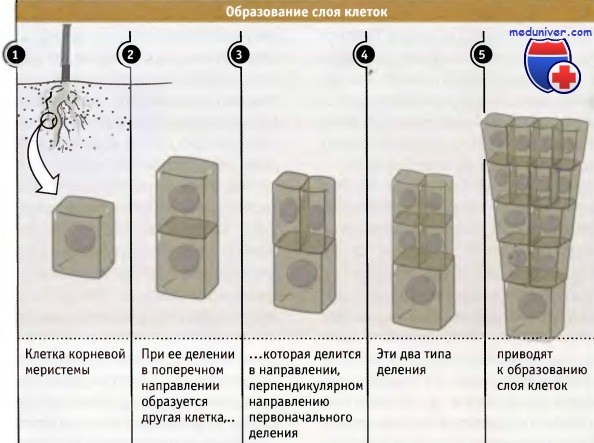
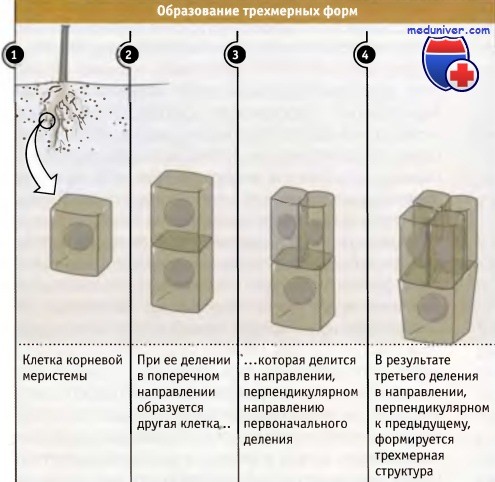
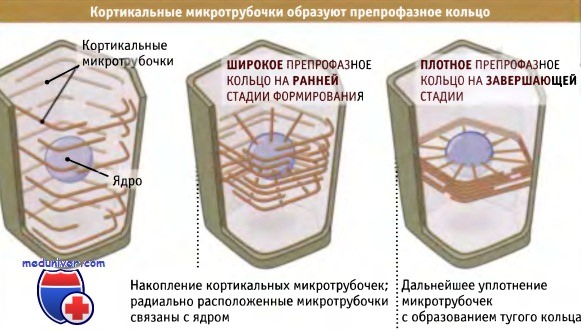
Направление плоскости деления клеток растений и его значение• При отсутствии у клеток подвижности, ориентация плоскости деления помогает определить форму • Формирующее деление (деление, способствующее развитию) приводит к появлению нового типа клеток; пролиферативное деление увеличивает их количество Наличие стенки у каждой клетки растения имеет важные последствия для их развития и формообразования. В отличие от клеток млекопитающих, которые могут менять форму и локализацию, тем самым изменяя строение тела, клетки меристемы растений неподвижны. Их стенки сохраняют определенную форму, связывая клетки между собой таким образом, что они не могут взаимно сдвигаться. Поэтому для клеток растений деление представляет собой единственную возможность их определенной ориентации друг относительно друга. Присутствие клеточной стенки означает, что деление не может происходить за счет сокращения клеточного кортекса, как это наблюдается при цитокинезе клеток животных. Вместо этого клетки растений делятся, образуя внутриклеточные перегородки. Направление плоскостей этих поперечных перегородок тщательно контролируется, поскольку, если такая перегородка образовалась, она не может быть повернута или передвинута в новое положение. Обеспечение ориентировки клеточного деления в меристеме является важнейшим процессом, который используется для того, чтобы определенным образом расположить клетки в растении и организовать их так, чтобы они были способны образовать при дифференцировке функционирующую ткань. Для того чтобы понять, как изменения направления плоскости деления влияют на образующуюся ткань, рассмотрим простой пример деления клетки, имеющей прямоугольную форму, показанный на рисунке ниже. Если исходная и все дочерние клетки делятся в одинаковом поперечном направлении, то образуется ряд клеток, связанных своими концами. Расширение в одном направлении может вызвать растяжение этого ряда; однако, поскольку все клетки в нем продолжают делиться в той же ориентации, ряд будет сохранять свою структуру. Такая форма роста характерна для простейших растений, например для нитчатых водорослей.
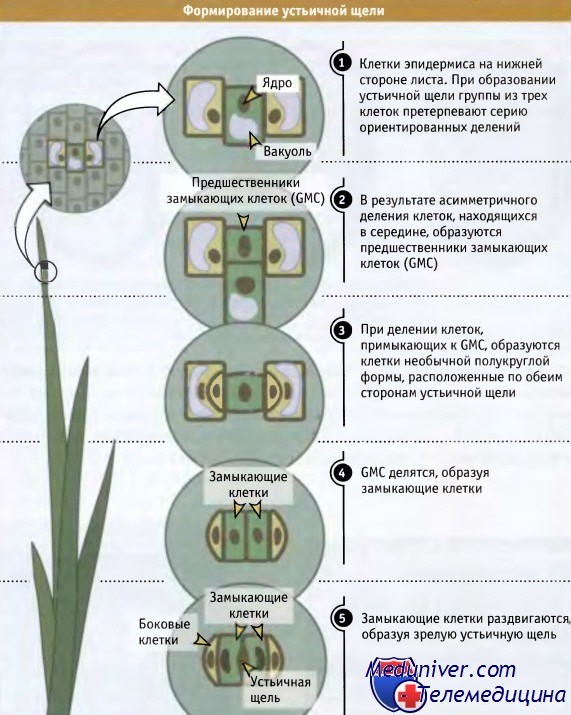
Однако, если в некоторой точке одна из клеток разделится перпендикулярно относительно плоскости предыдущего деления, один ряд окажется расщепленным на два параллельных. Повторное расщепление рядов в этой второй плоскости образует слой клеток, напоминающий характерный для поверхности листа. Конечно, растение может расти в трех направлениях, и определенная последовательность делений, скоординированная во времени с их ориентацией, дает возможность образоваться трехмерным формам. Формирование корня в трех направлениях требует скоординированности различного типа делений. Корень можно представить как состоящий из рядов клеток, собранных в пучки. Как обсуждалось выше, в ряду клетка может делиться или в направлении, перпендикулярном оси ряда, или вдоль него. Такие деления называются или поперечными, или продольными. Каждый тип деления вносит свой вклад в формирование корня, и его окончательные размеры определяются локализацией и количеством делений каждого типа. Поперечное (или «пролиферативное») деление увеличивает длину корня за счет увеличения в ряду количества одного определенного типа клеток. Если мы возьмем индивидуальный ряд, который расположен гораздо ниже меристемы, и проследим его путь по направлению к кончику корня, то сможем увидеть, что сам ряд произошел в результате по-разному ориентированных делений специальной базальной клетки. Такие деления создают новые ряды клеток в продольном направлении и называются «формирующими» делениями. Две дочерних клетки и их потомки, которые образуются в результате формирующего деления, могут иметь разную судьбу. Таким образом, независимо от ткани, каждый ряд клеток создается в ней задолго до наступления дифференцировки за счет продольного деления базальной клетки, расположенной вблизи кончика корня. Такая трехмерность строения хорошо видна на поперечных срезах корней Arabidopsis thaliana, небольшого растения, которое широко используется для генетических экспериментов. На срезах видно стереотипическое расположение клеток, которые собраны в концентрические слои, причем каждый слой состоит из строго определенного количества клеток Роль формирующего деления в создании такого расположения клеток иллюстрируется свойствами мутантов, у которых не образуется концентрических слоев и не формируется нормальная структура корня. Тот факт, что характер расположения клеток у различных растений одинаков, позволяет предполагать, что последовательность ориентированных делений, в результате которых она создается, хорошо воспроизводится и, вероятно, находится под генетическим контролем. Известны, однако, случаи, когда направление плоскости клеточного деления играет не столь критическую роль в формообразовании растений. Иногда, например, в листе, вместо обычного поперечного деления, в результате которого ряды клеток удлиняются, клетки начинают делиться под углом к каждому ряду. В результате клетки располагаются по-другому, хотя общая форма листа при этом сохраняется. Иногда в результате ориентированных клеточных делений образуются небольшие группы клеток и формируются органы или возникают клетки специфической формы. Это происходит при образовании устьиц, или мельчайших пор на нижней поверхности листа, которые открываются и закрываются, регулируя доступ газов и воды внутрь. Устьица состоят из небольшого количества клеток: они содержат разные количества (в зависимости от вида растения) «вспомогательных» клеток и две «сторожевых клетки» крестообразной формы, которые обладают подвижностью за счет изменения тургора и способны открывать и закрывать пору, находящуюся между ними. Эта группа клеток и пора образуется за счет серии ориентированных клеточных делений, происходящих в определенной последовательности, как показано на рисунке ниже. Процесс начинается с поперечного деления клеток в эпидермальном ряду; деление носит асимметричный характер, и образующаяся клетка называется Сторожевая материнская клетка, из которой возникают сторожевые клетки. Эта клетка представляет собой меньшую из двух клеток. Соседние клетки, находящиеся в двух примыкающих рядах, также делятся асимметрически, образуя клетки, которые покрывают сторожевую материнскую клетку. Последняя затем делится симметрично, и в продольном направлении, параллельно ряду клеток. При этом формируются две сторожевых клетки, которые затем отделяются друг от друга, образуя пору устьица. Таким образом, в функционировании этой структуры существенную роль играет образование различных плоскостей направления деления, которые обеспечивают возникновение комплекса устьица.
— Также рекомендуем «Формирование направления плоскости деления клеток растений» Оглавление темы «Цитология растений»:
|
Источник